

Глава 4. Головной мозг и глаз
 ральнои
медленной стадии и ипсилатеральнои
быстрой.
ральнои
медленной стадии и ипсилатеральнои
быстрой.
Для вертикальных движений глаза вестибулярные нейроны проецируются через медиальный продольный пучок (и, возможно, соединительное плечо). Но эксцитатный (возбуждающий) путь, идущий от переднего канала до ипсилатеральнои верхней прямой мышцы и контрлатеральных нижних косых мышц, проходит в верхнюю мозжечковую ножку (соединительное плечо).
При стимуляции передних и задних полукружных каналов вестибулярная система формирует «медленный» компонент вертикального нистагма. Особенностью является то, что в вертикальных движениях участвуют верхние и медиальные вестибулярные ядра. При этом стимулируются как эксцитатные, так и ингиби-рующие пути.
Необходимо обратить внимание на то, что произвольный взгляд и вестибулярные механизмы конкурируют в парамедианной ретикулярной формации моста. При этом произвольный взгляд ингибирует вестибулярный нистагм.
Вестибуло-глазной рефлекс обладает большой пластичностью. В этом процессе большую роль играет мозжечок, изменяющий силу обратной связи [372, 444]. При этом мозжечок использует зрительную информацию, получаемую от сетчатки во время движения туловища,
с целью определения «силы обратной связи». Именно благодаря этой информации мозжечок формирует «сигнал ошибки», который и используется для увеличения или уменьшения «силы обратной связи».
Оптокинетическая система. Когда испытуемый фиксирует равномерно движущийся зрительный стимул, происходит периодическое чередование саккад и медленных следящих движений глаз. Это и есть оптокинетический нистагм. Развивается он при стимуляции периферии сетчатки [17, 96, 201].
Подобно вестибуло-глазному рефлексу, оптокинетическая система активирует мышцы глаза для стабилизации взора в течение вращения головы. Однако она отличается тем, что использует зрительную информацию. Как и вестибуло-глазной рефлекс, оптокинетический нистагм обладает «силой обратной связи», которую можно измерить. Этот показатель представляет собой отношение скорости вращения глаза к скорости вращения изображения.
В процессе формирования оптокинетического нистагма зрительные афференты проецируются на вестибулярные ядра несколькими путями. Это облегчает интеграцию вес-тибуло-глазного и оптокинетического рефлексов. Схема, иллюстрирующая возможные пути оптокинетической системы, представлена на рис. 4.4.26, 4.4.28.
 Горизонтальная
Горизонтальная
Поворот глаз в противоположную сторону в горизонтальной плоскости (медленная фаза нистагма)
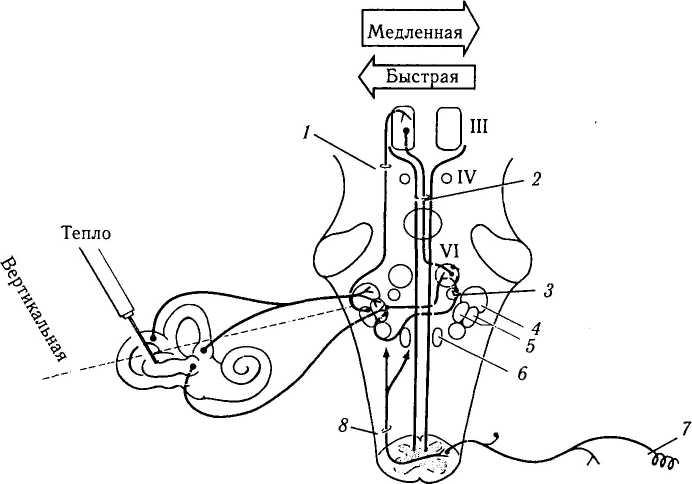
Рис. 4.4.26. Надъядерный контроль вестибулярной и проприорецептивной рефлекторных систем:
/ — восходящий тракт Дейтерса (эксцитатный); 2 — медиальный продольный пучок (эксцитатный); 3 — парамедианная ретикулярная формация моста (эксцитатная); 4 — верхнее вестибулярное ядро; 5 — медиальное и латеральное вестибулярные ядра; 6 — п. prepositus hypoglossi; 7 — рецептор мышцы; 8 — спин-но-вестибулярный тракт. Эксцитатный тракт выделен красным цветом. Нервные волокна, исходящие из горизонтального полукружного канала, направляются к латеральному вестибулярному ядру. Во время медленной фазы нистагма интернейроны вестибулярного ядра возбуждают контрлатеральную парамедианную
ретикулярную формацию моста. В свою очередь, возбуждаются контрлатеральные нейроны отводящего нерва и субъядро ипси-латеральной внутренней прямой мышцы, благодаря медиальному продольному пучку. Во время быстрой фазы нистагма часть ин-гибирующих импульсов идет к ипсилатеальному субъядру внутренней прямой мышцы через восходящий тракт Дейтерса, который располагается латеральней медиального продольного пучка. Контрлатеральные проприорецептивные рецепторы и восходящие спинно-вестибулярные волокна также оканчиваются вблизи вестибулярных ядер и п. prepositus hypoglossi
Движения глаз
517
 Вращение
туловища стимулирует оба рефлекса
следующим образом: сначала развивается
вестибуло-глазной рефлекс [402, 372] и
затем
подкрепляется оптокинетическим
рефлексом.
В течение более длительного вращения
с открытыми глазами вестибулярный
компонент постепенно
прекращается, поскольку прекращается
движение эндолимфы. Оптокинетический
нистагм при этом сохраняется, поскольку
сохраняется
активность нейронов вестибулярных
ядер.
Вращение
туловища стимулирует оба рефлекса
следующим образом: сначала развивается
вестибуло-глазной рефлекс [402, 372] и
затем
подкрепляется оптокинетическим
рефлексом.
В течение более длительного вращения
с открытыми глазами вестибулярный
компонент постепенно
прекращается, поскольку прекращается
движение эндолимфы. Оптокинетический
нистагм при этом сохраняется, поскольку
сохраняется
активность нейронов вестибулярных
ядер.
НКТв
НКТд
Структурно оптокинетический нистагм обеспечивается многими образованиями мозга. В его формировании принимают участие: лобно-ви-сочная кора, задняя часть теменной коры, пре-центральная и задняя срединная лобная извилины, передний и задний островок, прелобная кора и средняя часть верхней лобной извилины. К подкорковым структурам, которые активизируются при оптокинетическом нистагме, относятся хвостатое ядро, бледное ядро (globus pallidus) и парамедиальная часть таламуса [72, 145, 236, 272, 389]. При оптокинетическом нистагме информация, получаемая сетчаткой, достигает также вестибулярных ядер через добавочный зрительный тракт, включающий в свой состав ядро зрительного пути [96, 117, 201] (рис. 4.4.18, 4.4.27), ретикулярные ядра покрышки моста (nuclei tegmenti) [271, 299], а также медиальное ядро моста (nucleus pontis) [300]. В процесс вовлечены также п. perihypo-glossi [557] и вестибулярная спайка [139].
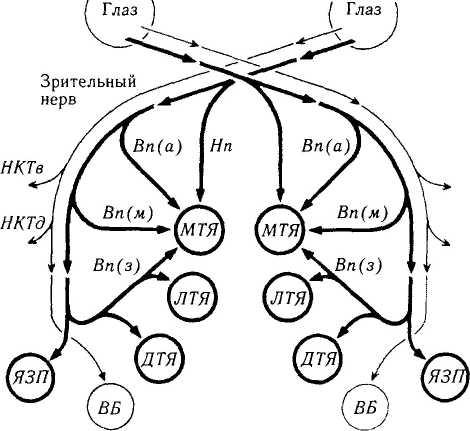
Рис. 4.4.27. Схематическое изображение состава и связей ядер дополнительной системы зрительного тракта (по Lenerstrand et ai, 1982):
НКТв — вентральное наружное коленчатое тело; НКТд — дор-зальное наружное коленчатое тело; МТЯ — медиальное терминальное ядро; ЛТЯ — латеральное терминальное ядро; ДТЯ — дорзальное терминальное ядро; ВБ — верхние бугорки четверохолмия; ЯЗП — ядро зрительного пути; Нп — нижний пучок, Вп(а) — верхняя часть верхнего пучка, Вп(м) — медиальная часть верхнего пучка; Вп(з) — задняя часть верхнего пучка
Значение коры в оптокинетическом нистагме поддерживается экспериментальными исследованиями. Так, двусторонняя лобэктомия, проведенная у обезьян, вызывает потерю плавного компонента слежения оптокинетического нистагма, а повреждение медиальной верхней височной области (поля 19 и 39) ведет к нарушению следящих движений глаза в сторону повреждения [550].
У людей с односторонними теменными поражениями выявляется асимметрия плавного слежения [112, 166].
Выше неоднократно подчеркивалось большое значение мозжечка в регуляции движений глаза. В чем же оно состоит и как осуществляется? Необходимо отметить, что мозжечок только коррегирует движения. Осуществляется это сложной системой связей его со многими структурами мозга. Перед тем как остановиться на этих связях, необходимо кратко напомнить строение мозжечка, частично описанное в первом разделе этой главы (рис. 4.1.28, 4.1.29).
У высших млекопитающих мозжечок состоит из нового мозжечка, или неоцеребеллума (полушария и участки червя), старого, или па-леоцеребеллума (участки червя, соответствующие передней доле, пирамиды, язычок и пара-флокулярные отделы), и древнего мозжечка, или архицеребеллума (клочково-узелковая доля; lobus flocculonodularis (рис. 4.1.28, 4.1.29, 4.4.29).
Эта классификация соответствует его подразделению в зависимости от поступающих афферентных путей. По этой причине архицере-беллум называют вестибулоцеребеллум, палео-церебеллум — спиноцеребеллум, а неоцеребел-лум — понтоцеребеллум.
В белом веществе мозжечка заключено несколько ядер — ядро шатра (п. fastigii), шаровидное (п. globosus), пробковидное (п. emboli-formis) и зубчатое (п. dentatus) ядра.
Сложной структурой обладает кора мозжечка (рис. 4.4.30, см. цв. вкл.). В различных слоях коры расположены шесть типов нервных клеток и их отростки. Мелкие клетки-зерна лежат в зернистом слое, а аксоны их идут в молекулярный слой. Здесь эти аксоны Т-образно делятся, посылая в каждом направлении вдоль поверхности коры ветвь (параллельное волокно) длиной 1—2 мм. Эти ветви проходят через области ветвления остальных типов нейронов и образуют на них синапсы. В зернистом слое расположены клетки Гольджи, дендриты которых распространяются в молекулярном слое, а их аксоны направляются к клеткам-зернам.
Между этими двумя слоями лежит слой нейронов Пуркине, дендриты которых разветвляются в молекулярном слое. Аксоны клеток спускаются к ядрам мозжечка, и небольшое их количество заканчивается в вестибулярных ядрах.
Остальные три типа клеток — корзинчатые, звездчатые и клетки Лугаро — лежат в моле-
518