

Глава 4. Головной мозг и глаз
 к
бесконечному числу возможных вариантов
оттенков
цвета, наиболее логично предположить
существование трех основных рецепторов,
воспринимающих
красный, синий и зеленый цвета.
Последующие исследования Гельмгольца
и Максвелла
показали, что психофизиологические
реакции человека на цветовые стимулы
наиболее
полно можно описать существованием
рецепторов, реагирующих на три различные
длины волны. Лишь в XX
столетии
благодаря разработке
метода микроспектрофотометрии были
выделены три типа колбочек, каждый из
которых
содержал особый пигмент, обусловливающий
избирательность по отношению к цвету.
Различные пигменты по-разному поглощают
свет,
обладая избирательностью по отношению
к
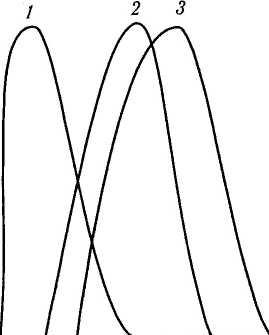
определенным длинам волн (рис. 4.2.13).
Один
класс зрительных пигментов имеет
спектральную
чувствительность, которая достигает
максимума
от 445 до 450 нм,
другой
— от 525 до
535 нм;
максимум
поглощения третьего пигмента лежит
в области 555—570 нм.
Трем
пигментам
соответствуют три типа рецепторов,
обусловливающих
нормальное цветовое зрение человека.
Из описания чувствительности этих
первичных
рецепторов можно вывести почти все
феномены цветового зрения человека.
Например,
из концепции круга следует, что смесь
зеленого
(520 нм)
и
красного (620 нм)
должна
быть
неотличима от желтого с длиной волны
564
нм.
к
бесконечному числу возможных вариантов
оттенков
цвета, наиболее логично предположить
существование трех основных рецепторов,
воспринимающих
красный, синий и зеленый цвета.
Последующие исследования Гельмгольца
и Максвелла
показали, что психофизиологические
реакции человека на цветовые стимулы
наиболее
полно можно описать существованием
рецепторов, реагирующих на три различные
длины волны. Лишь в XX
столетии
благодаря разработке
метода микроспектрофотометрии были
выделены три типа колбочек, каждый из
которых
содержал особый пигмент, обусловливающий
избирательность по отношению к цвету.
Различные пигменты по-разному поглощают
свет,
обладая избирательностью по отношению
к
определенным длинам волн (рис. 4.2.13).
Один
класс зрительных пигментов имеет
спектральную
чувствительность, которая достигает
максимума
от 445 до 450 нм,
другой
— от 525 до
535 нм;
максимум
поглощения третьего пигмента лежит
в области 555—570 нм.
Трем
пигментам
соответствуют три типа рецепторов,
обусловливающих
нормальное цветовое зрение человека.
Из описания чувствительности этих
первичных
рецепторов можно вывести почти все
феномены цветового зрения человека.
Например,
из концепции круга следует, что смесь
зеленого
(520 нм)
и
красного (620 нм)
должна
быть
неотличима от желтого с длиной волны
564
нм.
Необходимо отметить, что, используя цветовой круг, возможно описать и цвета последовательных образов. Напомним, что последовательный образ возникает в тех случаях, когда наблюдатель длительно смотрит на поверхность одного цвета, а затем переводит взгляд на белую поверхность. При этом возникает восприятие цвета, соответствующего дополнительному цвету, на который ранее смотрел наблюдатель. Цвет последовательного образа в цве-
100
75
§1
5о
!
S
я о
§•
25
300 400 500 600
Длина волны, нм
700
Рис. 4.2.13. Кривые поглощения световой энергии «синими» (/), «зелеными» (2) и «красными» (3) колбочками
товом круге лежит напротив цвета, который первоначально наблюдался наблюдателем. Последовательные образы подчиняются правилу взаимности. Если синий цвет дает последовательный образ желтого цвета, то желтый дает последовательный образ синего цвета. Приведенные выше наблюдения явились основой создания теории оппонентных процессов в цветовом зрении, которую предложил еще в 1878 г. Эвальд Геринг, а развил Харинг.
В соответствии с этой теорией цвета смешиваются по парам дополнительных цветов: синий с желтым, красный с зеленым и белый с черным (рис. 4.2.11, б). Оценка яркости выводится на основе реакции механизма «черный — белый», а цвета возникают из комбинаций механизмов «синий — желтый» и «красный — зеленый».
Согласно одной из предложенных схем, три основных рецептора цвета присоединены к трем нервным клеткам так, что получаются пары «красный — зеленый», «черный — белый» и «синий — желтый». Факт образования различных сочетаний рецепторами доказан нейрофизиологами и нейрогистологами, на чем мы остановимся несколько ниже.
При рассмотрении цветовосприятия необходимо описать еще одно явление, а именно, явление наведенного контраста. При восприятии цвета, так же как и черно-белых объектов, восприятие данного участка влияет на восприятие соседних участков. Это связано с латеральным торможением, т. е. с тем же явлением, которое обусловливает усиление контраста и константность яркости. Если смотреть на синее пятно в данном участке, то при этом снижается чувствительность к синему в соседних участках и, следовательно, повышается чувствительность к желтому. Таким же образом восприятие белого цвета повышает чувствительность к черному; восприятие красного повышает чувствительность к зеленому цвету. Такие контрастные эффекты носят название пространственного, или индуцированного, контраста: каждый данный цвет наводит на соседние участки поля цвет, являющийся для него дополнительным.
Для того чтобы объяснить наблюдения такого рода, надо допустить, что один основной нейрон, приходя в состояние возбуждения, тормозит все окружающие нейроны того же класса. Поскольку эти рецепторы спарены и образуют оппонентные системы, угнетение одного рецептора равносильно повышению чувствительности его оппонента. В этом причина цветовых контрастов. Наиболее выражен эффект контраста в парах дополнительных цветов. В результате деятельности оппонентной системы можно получить почти полный набор воспринимаемых цветовых тонов.
Необходимо отметить, что на протяжении многих десятилетий две приведенные выше теории цветового восприятия (трехцветная
Функциональная анатомия зрительной системы
423

 и
оппонентная) рассматривались как
исключающие
одна другую. В настоящее время стало
ясно,
что как трехкомпонентная теория, так
и оппонентная
теория пригодны для описания механизмов
цветового восприятия. Причем первая
достаточно четко описывает цветорецеп-цию
на уровне фоторецепторов, а вторая
обеспечивает
обработку цветовой информации на более
высоких уровнях зрительной системы
(наружные
коленчатые тела, кора большого мозга).
В последние годы в экспериментах на
обезьянах
выявлено, что функционирование
оппонентного
механизма цветовосприятия реализуется
не только на уровне наружного коленчатого
тела и коры головного мозга, но и на
уровне сетчатки [130, 363].
и
оппонентная) рассматривались как
исключающие
одна другую. В настоящее время стало
ясно,
что как трехкомпонентная теория, так
и оппонентная
теория пригодны для описания механизмов
цветового восприятия. Причем первая
достаточно четко описывает цветорецеп-цию
на уровне фоторецепторов, а вторая
обеспечивает
обработку цветовой информации на более
высоких уровнях зрительной системы
(наружные
коленчатые тела, кора большого мозга).
В последние годы в экспериментах на
обезьянах
выявлено, что функционирование
оппонентного
механизма цветовосприятия реализуется
не только на уровне наружного коленчатого
тела и коры головного мозга, но и на
уровне сетчатки [130, 363].
Теперь мы более подробно остановимся на структурном обеспечении восприятия цвета зрительной системой. Как было указано выше, колбочки человека и приматов отличаются пиками чувствительности к разным длинам видимой части электромагнитного излучения. Это так называемые коротковолновые колбочки (S или «синие колбочки»), колбочки, реагирующие на среднюю длину волны (М или «зеленые колобочки»), и длинноволновые колбочки (L или «красные колбочки»). Большая часть ганглиозных клеток сетчатки отличается цвето-оппонентной реакцией на цвет (60% нейронов у обезьян). Они возбуждаются при освещении сетчатки одной длиной волны и тормозятся другой. «Красно-зеленые» ганглиозные клетки формируют оппонентный ответ путем взаимодействия сигналов, идущих от М- против L-колбочек. «Сине-желтые» клетки формируют оппонентный ответ путем взаимодействия сигналов, поступающих от S и определенной комбинации М- и L-колбочек (рис. 4.2.14—4.2.17, см. цв. вкл.).
В последние годы были выявлены механизмы обеспечения колбочковой оппоненции благодаря разработке и использованию ряда новых методов исследования.
Во-первых, изучение сетчатки обезьяны in vitro выявило, что особенности оппонентного ответа связаны с различным морфологическим типом нейронов сетчатки. Это стало возможным благодаря использованию переживающей культуры комплекса структур глаза, а именно — сосудистой оболочки и сетчатки. Сохранение анатомических соотношений указанных структур поддерживает специфический метаболизм нейронов сетчатки и позволяет снять потенциал отдельного нейрона. Этот метод позволил также подводить световую энергию различных спектральных характеристик к отельному фоторецептору, используя при этом довольно сложные оптические системы. Снятие потенциала нейронов сетчатки при этом сопровождалось микроскопическим исследованием сетчатки.
Во-вторых, установлено, что особенности организации оппонентных рецептивных полей связаны с характером связей между нейронами
и типом используемого нейроном нейромедиа-тора. Это стало возможным благодаря использованию иммуногистохимических и ультраструктурных исследований.
В-третьих, появилась возможность получить и математически обрабатывать нервный сигнал после снятия потенциала изолированного нейрона на разных уровнях зрительного анализатора при помощи микроэлектродов.
Использование выше приведенных современных морфологических методов исследования выявило, что большая часть ганглиозных клеток сетчатки относится к так называемым «карликовым» и «зонтикоподобными» клетками [195, 196, 314, 363, 422, 452, 605]. Именно по этой причине «цветовой тракт» назван карликовым.
В настоящее время принято объединение карликовых и зонтикоподобных ганглиозных клеток в единую группу так называемых Р-кле-ток. Эти клетки получили такое название по той причине, что они проецируются на парво-целлюлярные (мелкоклеточные) слои наружного коленчатого тела [133, 363, 503]. Существуют также ганглиозные клетки, проецирующиеся на магноцеллюлярные слои наружного коленчатого тела (М-тракт).
Разделение трактов на Р- и М-тракты имеет большое функциональное значение, поскольку между ними существуют большие различия, сводящиеся к следующему:
Р-клетки чувствительны к цветовому сти мулу, а М-клетки — нет.
М-клетки намного более чувствительны к черно-белым стимулам низкой интенсивности.
Рецептивные поля Р-клеток значительно меньше, чем Af-клеток.
Аксоны М-клеток проводят импульс зна чительно быстрее.
Реакция Р-клеток на зрительный стимул, особенно цветовой, довольно длительная, в то время как Af-клетки — кратковременная.
Таким образом, наиболее существенным отличием является то, что Р-тракт обеспечивает цветовую чувствительность, а Л1-тракт — контрастность.
Ранее предполагали, что и «сине-желтые» и «красно-зеленые» оппонентные ганглиозные клетки по морфологическим особенностям относятся к карликовым. Однако использование регистрации потенциала отдельной ганглиоз-ной клетки позволило выяснить, что нейроны, отличающиеся колбочковой цветооппоненцией, можно разделить на два морфологических типа. «Красно-зеленые» оппонентные клетки относятся к карликовым, а подгруппа «сине-желтых» оппонентных клеток («синий ON») — к бистратифицированным. Бистратифицирован-ные ганглиозные клетки являются единственными ганглиозными клетками, которые получают информацию от S-колбочек (рис. 4.2.14— 4.2.17).
424