

Глава 4. Головной мозг и глаз

типа
довольно хорошо определены. В то
же головного
мозга обезьяны (темные полосы), выявляе-
время
их роль в зрительном восприятии
пока мые
после введения в глазное яблока
радиоактивного
не
полностью ясна. Поскольку «простые»
и пролина:
«комплексные» клетки реагируют лучше
всего
r rJ J в
верхнем правом углу иллюстрируется
макроскопический вид
На
Ориентируемые В Пространстве КОНТурЫ
Зри- радиоавтографа
зрительной коры

|
д + л +■* л ++ i ♦ |
|
| |||||
|
|
|
|
| ||||
|
|
|
л |
|
| |||
Р
 ис.
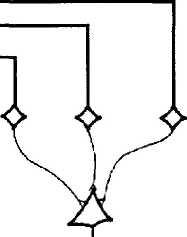
4.2.54. Схематические изображения,
иллюстрирующие
гипотезу Hubel,
Weisel (1962)
относительно организации
простых (а) и комплексных (б) рецептивных
полей:
ис.
4.2.54. Схематические изображения,
иллюстрирующие
гипотезу Hubel,
Weisel (1962)
относительно организации
простых (а) и комплексных (б) рецептивных
полей:
а — большое количество нейронов наружного коленчатого тела, четыре из которых представлено справа, обладает рецептивным полем с ON-центром, расположенным вдоль плоскости сетчатки. Все они проецируются на «простую клетку» коры и синапсы становятся эксцитатными. Рецептивное поле кортикальной клетки, в результате этого будет обладать продолговатым рецептивным полем с ON-центром, показанным слева; б — определенное количество клеток с простым рецептивным полем проецируется на одну «комплексную клетку». В этом случае «простая клетка» обладает продолговатым рецептивным полем, содержащим эксци-татные и тормозные зоны. Граница поля ограничена пунктирной линией. Последовательная активация вертикального края движущимся стимулом поперек этого прямоугольника, независимо от его положения, будет активировать «простые клетки» и вызывать эксцитатный ответ более высоко расположенного нейрона
ного мозга. Это находит свое отражение и в характере распределения нейронов с цветовыми рецептивными полями по слоям коры. Если клетки с «концентрическими» и «простыми» рецептивными полями обнаруживаются в III и IV слоях коры, тесно связанных с входами наружного коленчатого тела, то клетки со «сложными» и «сверхсложными» рецептивными полями проецируются на верхние (II и верхняя часть III слоя) и нижние (V и VI) слои коры. Из выше приведенных данных видно, что сейчас свойства рецептивных полей различного
тельного объекта, предполагают, что они обрабатывают информацию относительно границ или граней. Тем не менее не ясно, благодаря каким нейронам и на каком уровне происходит формирование «зрительного восприятия» как высшей формы обработки зрительной информации.
Структурно-функциональная организация первичной зрительной коры
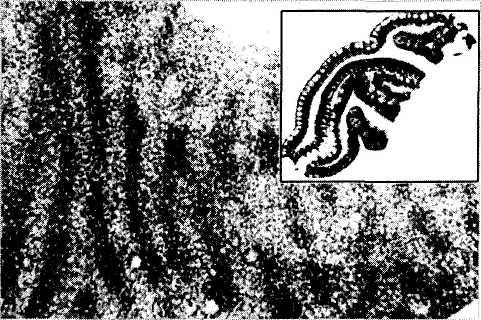
Глазные доминантные колонки. После выявления основных физиологических закономерностей реакции зрительной коры головного мозга на «световые стимулы» возник вопрос — какова структурная организация коры, обеспечивающая эти физиологические особенности? Решению этих вопросов содействовало применение новых методов исследования. На протяжении последних двух десятилетий нейронную структуру и функцию зрительной коры интенсивно изучали путем маркировки аксонов нейронов ганглиозных клеток сетчатки и наружного коленчатого тела. При исследовании терминальной дегенерации использовали методы радиоаутографии, а ретроградной дегенерации — введение пероксидазы хрена [256—270, 326, 333, 415]. Большинство исследований проведено на кошках и обезьянах. Показано, что многие закономерности организации коры, выявленные у этих животных, распространяются и на человека.
Рис. 4.2.55. Доминантные колонки зрительной коры
В настоящее время ясно, что в структурно-функциональном отношении нейроны первичной зрительной коры организованы в так называемые «колоночные единицы» [540] (рис. 4.2.50, 4.2.55, 4.2.56). Анатомическая реальность «колоночной» цитоархитектоники зрительной коры подтверждена методами марки-
Функциональная анатомия зрительной системы
453

Зрительное
поле (Y2)
Стриарная
кора (VI)
Верхние
бугорки четверохолмия
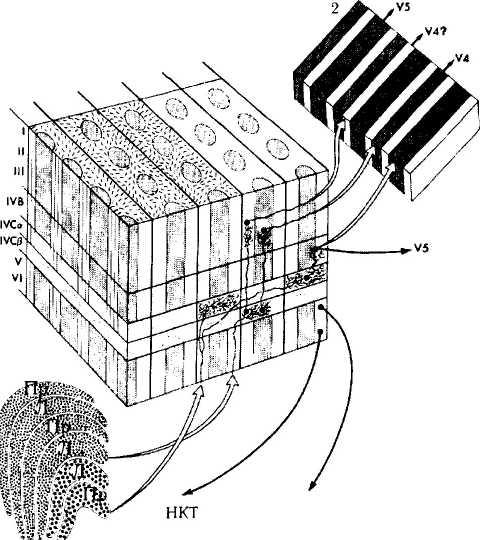
Рис. 4.2.56. Схематическое изображение магно- (М) и парвоцеллюлярных (Р) трактов, идущих от наружного коленчатого тела через поля VI и V2 к полям V4 и V5:
каждый модуль стриарной коры содержит определенное количество глазных доминантных колонок, ориентационных колонок и дающих положительную реакцию при выявлении активности ци-тохромоксидазы «капель» (заштрихованные цилиндры, которые прерываются в слое IVC). Ориентационные колонки (отмечены наклонными линиями на поверхности коры) распространяются через все слои, за исключением слоя IVB-бета. Магноцеллюляр-ный путь проходит через слои IVC-альфа, IVB и направляется к полям коры V2 и V5. Парвоцеллюлярный путь проецируется через слой IVC-бета на слои II и III. Клетки цитохромоксидазных капель проецируются на тонкую темную полосу в поле V2. Клетки, расположенные между капель, проецируются на светлые тонкие полосы V2, а затем, возможно, на V4. Слои V и VI проецируются на верхние бугорки четверохолмия и наружное коленчатое тело соответственно
ровки нейронов индикаторами типа радиоактивного пролина. После введения радиоактивного пролина внутрь глаза он распространяется по направлению наружного коленчатого тела, а затем через синапсы к первичной зрительной коре [259]. При использовании этого метода зрительная кора неравномерно окрашивается в виде «колонок». У человека «колонки» выявляются при определении активности цито-хромоксидазы.
Благодаря использованию подобных методов установлено, что нервные клетки зрительной коры распространяются не только по слоям, параллельным поверхности коры, но также существуют функциональные слои, расположенные перпендикулярно поверхности коры. Подобные структурные образования отсутствуют в месте проекции слепого пятна и «назального полумесяца».
Электрофизиологическая регистрация сигнала от нейронов IVC-слоя зрительной коры по-
зволила установить, что информация от наружного коленчатого тела направляется к смежным «колонкам», относящимся к правому и левому глазам, чередуясь при этом (рис. 4.2.50, 4.2.56). Подобные отношения структурно обеспечивают бинокулярные взаимоотношения на уровне коры. Монокулярно доминирующие «колонки» названы «глазодоминантными колонками». Именно расположением «глазодоминантных колонок» обоих глаз по соседству, а также наличием нейронных связей между ними обеспечивается «перекрывание» точек зрительного пространства, проецирующихся на левое и правое глазное яблоко.
Колоночные комплексы нейронов первичной зрительной коры большего размера, включающие в свой состав «глазодоминантные колонки» левого и правого глаза, а также, описанные ниже «ориентационные колонки», называются «гиперколонками». Площадь среза «гиперколонки» равняется примерно 1 мм2. На каждую «гиперколонку» проецируется определенная область зрительного поля. Причем размер «гиперколонки» зависит от участка сетчатой оболочки, проецирующейся на кору [348, 349, 451, 584, 607]. Наибольшим размером отличаются «гиперколонки», на которые проецируется периферия сетчатки, а наименьшим — маку-лярная область. Правда, необходимо отметить, что полного соответствия проекции сетчатой оболочки на зрительную кору не определяется, что является причиной дальнейших интенсивных исследований с использованием методов электрофизиологии [60]. Колончатой структурой обладают и детекторы цвета. В «цветовой колонке» нейроны реагируют только на монохроматическое излучение в узком диапазоне спектра. Каждая «колонка» детектирует специфическую полосу спектра. «Цветовые колонки» чередуются. «Колонки» с нейронами, селективно выделяющими область 490—500 нм, сменяются «колонками», выделяющими область 610 нм и т. д.
Ориентационные колонки и обработка информации. Помимо «глазодоминантных колонок» в зрительной коре выявлены и другие структурно-функциональные единицы, определяющие возможность определения пространственной ориентации зрительного объекта [65, 155]. Эти структуры были обнаружены благодаря использованию методов электрофизиологии. Впервые наличие ориентационной организации зрительной коры выявили Hubel и Wiesel [262, 263]. Вводя микроэлектрод строго перпендикулярно поверхности первичной зрительной коры, авторы установили, что все нейроны этой области отвечают только при одном условии — наличии определенной ориентации «светового сигнала» в виде щели или грани. Эти данные указывают на то, что эти нейроны сгруппированы в «колонки». Названы они «ориентацион-ными колонками» (рис. 4.2.57).
451