

Глава 4. Головной мозг и глаз

Вид с внутренней стороны
В настоящее время установлено, что в этих процессах, помимо обширных областей затылочной коры, участвуют также височная и теменная кора. Причем каждая из областей выполняет строго определенные функции (см. ниже). Самой важной закономерностью функционирования экстрастриарнои коры является иерархическая и параллельная обработка зрительной информации. Более того, экстра-стриарные области обладают ретинотопически-ми картами зрительного пространства. Известны и стереотопические проекции от одной зрительной области к другой. Эти связи, как правило, двухсторонние.
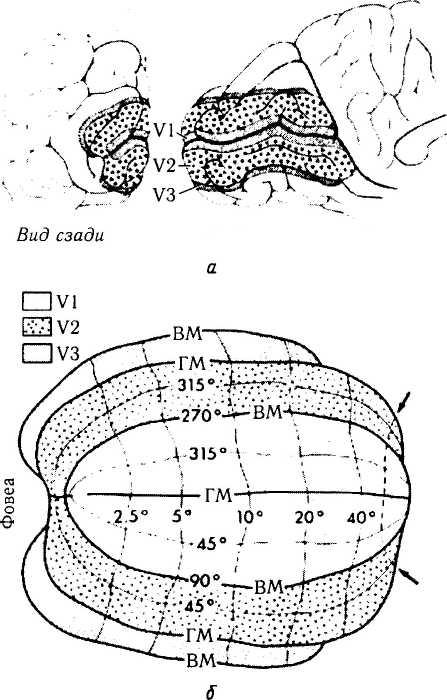
Первоначально мы охарактеризуем 18-е и 19-е поля.
Вторичная зрительная кора (поле 18) непосредственно прилежит к 17-му полю. Полоска Дженнери в ней отсутствует. На вторичную зрительную кору (кора нижней височной доли; поле 18), т. е. место, определяющее зрительную память, проецируется 17-е поле. Наличие подобной проекции объясняет механизм развития зрительной дезориентации, потери «топографической» памяти, зрительной агнозии и потерю
способности оценить расстояние до объекта при повреждении зрительной коры.
Третичная зрительная кора (поле 19) окружает вторичную кору на медиальной поверхности полушария мозга. Большая ее часть переходит на заднюю теменную долю, а снизу она формирует часть височной доли. При гистологическом исследовании третичная зрительная кора напоминает кору теменной доли, за исключением того, что в V слое отсутствуют большие пирамидные нейроны.
В срезах, окрашенных по Нисслю, эти поля выглядят довольно однородно и иногда даже квалифицированные анатомы не могут найти существенных различий между ними. Последние исследования с использованием электрофизиологических методов и радиоактивных трей-серов выявили в полях 18 и 19, по крайней мере, пять подобластей, отличающихся в функциональном отношении и принимающих участие в обработке зрительной информации. Это поля V2, V3, V3A, V4 и V5 [60, 77, 144] (рис. 4.2.59, 4.2.61, 4.2.62). Существует значительно больше полей, анализирующих зрительную информацию, но расположенных вне полей 18 и 19. К настоящему моменту у обезьян выявлено 25 подобных областей коры и семь ассоциативных областей, обеспечивающих связи с другими известными зрительными областями [322]. Эти области связаны между собой 305 отдельными проводящими путями. 32 зрительные области у обезьян занимают приблизительно 55% площади неокортекса. Это указывает на то, что обработка зрительной информации требует большого количества ткани коры мозга. Первичная зрительная кора в среднем составляет 1200 мм2, или приблизительно 12% неокортекса. Поле V2 несколько меньше и является второй по величине корковой областью. Вместе поля VI и V2 занимают примерно 20% полной площади поверхности неокортекса. Увеличение площади
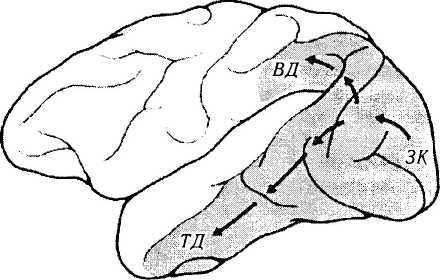
Рис. 4.2.61. Зрительные поля коры головного мозга обезьяны (по Mishkin, 1983):
стрелками указаны два кортикальных зрительных пути, начинающихся из первичной зрительной коры (ЗК). Первый путь направляется вентрально по направлению височной коры (ВД). Второй путь направляется дорзально в кору теменной доли (ТД). Вентральный путь является ключевым в объектном зрении, а дорзальный в пространственном
Функциональная анатомия зрительной системы
■157

КОРА
Лобное
глазное поле
(поле 8)
Нижняя
височная
(поля
20, 21)
I
1
| Задняя I
'
париетальная I
I 7
I
(поле
7)
V4
(поле
18)
V3a
(поле
18)
Престриарная
кора
Первичная
зрительная
кора
М
(магноцеллюлярный
тракт)
Р
(парвоцеллюлярный
тракт)
Парвоцеллюлярные
слои
Наружное
коленчатое
тело
СЕТЧАТКА
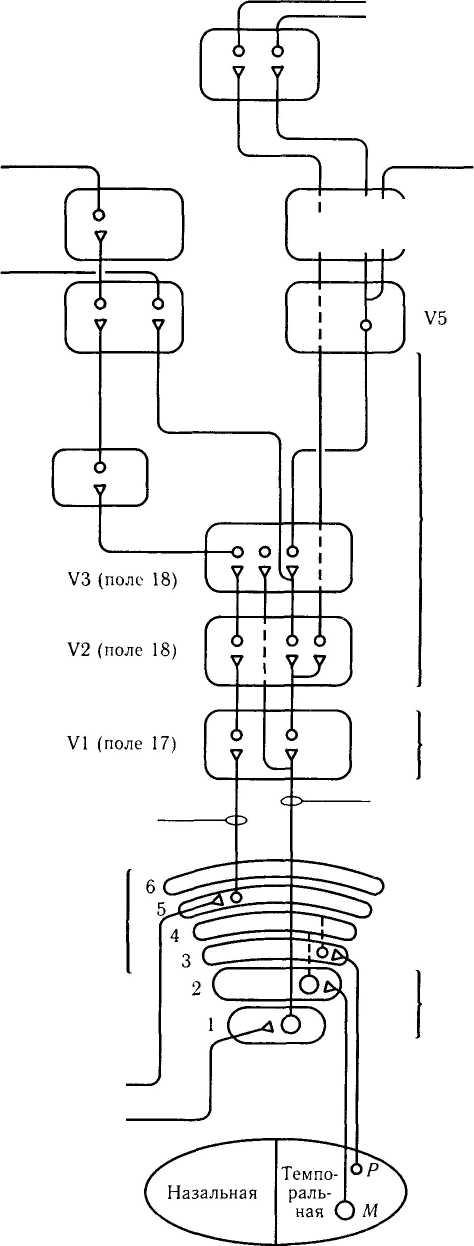
1985):
с одной стороны, зрительный путь характеризуется наличием довольно сложной иерархии структурных образований, идущих от сетчатой оболочки до различных участков коры головного мозга. С другой — существует два параллельных пути передачи зрительной информации — Р- и М-тракты. Парвоцеллюлярный (Я) тракт обеспечивает восприятие деталей, формы и цвета зрительного объекта, в то время как магноцеллюлярный (М) тракт анализирует движение цели
458