

Глава 4. Головной мозг и глаз
 Теперь
мы кратко охарактеризуем каждую из
указанных систем.
Теперь
мы кратко охарактеризуем каждую из
указанных систем.
Нейронная система палочек. Палочки и связанные с ними нейроны сетчатки ответственны за функционирование трактов, обеспечивающих «ночное зрение». То есть эта система нейронов, повышая чувствительность зрительной системы, позволяет анализировать зрительные объекты при низком освещении (скотопи-ческие условия).
Несмотря на то, что у человека существует «центральное зрение», определяющее высокую разрешающую способность зрительного анализатора, и цветовое зрение, система палочек также важна. При патологии нейронов этой системы развивается ночная слепота, наиболее часто встречающаяся при пигментном ретините.
Показано, что в сетчатке человека преобладают палочки, за исключением фовеолы. Их концентрация достигает максимума в кольце вокруг фовеа, шириной приблизительно 5 мм [405]. Высокая плотность фоторецепторов обеспечивает, в значительной степени, остроту зрения (см. главу 3). Максимум поглощения световой энергии пигментом палочки располагается в области 497 нм.
Каким же образом организована система палочек?
Сигнал, сформированный в момент освещения, от палочки передается биполярным клеткам. Выявляется только один морфологический тип биполярной клетки, который формирует си-наптическую связь с палочкой (синапс ленточ-
ного типа). Эта биполярная клетка одномоментно получает информацию от 15—30 палочек, что является морфологической основой конвергенции сигнала. Происходит это в наружном плексиформном слое (рис. 4.2.3; 4.2.8, см. цв. вкл.). Затем биполярные клетки палочек посылают аксоны в направлении внутреннего плексиформного слоя, где они заканчиваются в глубоких его слоях вблизи тел ганглиозных клеток. При этом прямого контакта между биполярной и ганглиозной клетками нет, а взаимодействие между ними осуществляется посредством амакриновых клеток. Вся обработка информации происходит на уровне дендритов амакриновых клеток (внутренний плексиформ-ный слой).
Функции большинства типов амакриновых клеток изучены пока недостаточно. Тем не менее известно, что они обеспечивают антагонистическое окружение рецептивного поля, благодаря механизму латерального торможения. Подобная организация нейронов обеспечивает дивергенцию и конвергенцию сигнала палочек на уровне биполярных клеток палочек и амакриновых клеток перед тем, как информация поступает ганглиозной клетке.
Формирование рецептивного поля осуществляется несколькими типами амакриновых клеток (All, A17).
Амакриновая клетка АН типа характеризуется тем, что при помощи щелевого соединения (электрический синапс) она соединяется с биполярной клеткой колбочки (рис. 4.2.8, 4.2.9).
Палочки
Колбочки
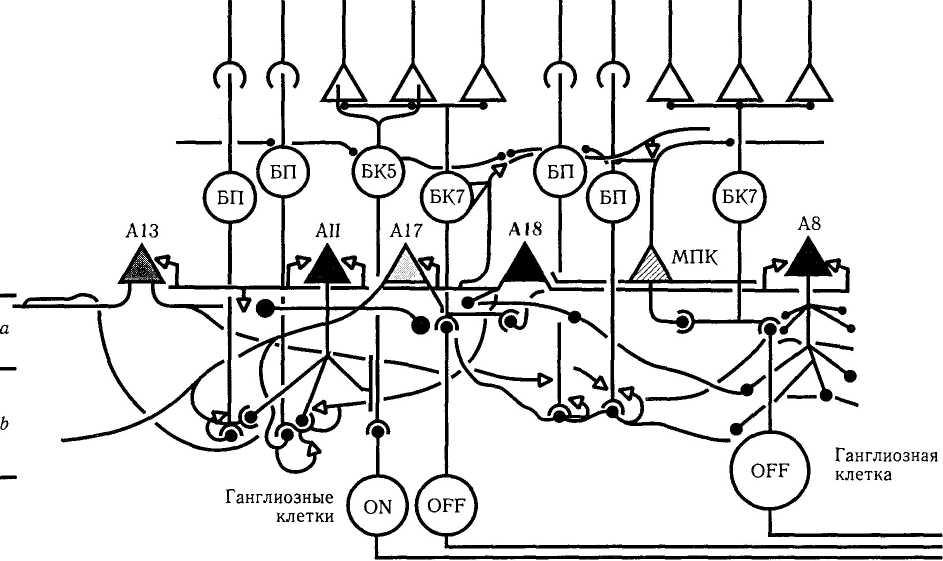
Рис. 4.2.9. Особенности связей амакриновой клетки типа А18 (объяснение в тексте)
Функциональная анатомия зрительной системы
419

 Последняя,
в свою очередь, вступает в контакт с
ганглиозной клеткой колбочки, что
происходит
в субслое b
внутреннего
плексиформного слоя
[314]. В результате этого формируется так
называемый «возвратный синапс»,
известный
как реципроктный синапс. Эти ганглиоз-ные
клетки отвечают на освещение
деполяризацией,
формируя ON-центр
рецептивного поля. В
то же время амакриновые клетки АН,
контактирующие
с ганглиозной клеткой при помощи
синаптической ленты в субслое а,
отвечают
на
освещение сетчатки гиперполяризацией,
образуя
OFF-центр
рецептивного поля (рис. 4.2.8, 4.2.9). Именно
благодаря этому образуются рецептивные
поля ON/OFF-типа.
Последняя,
в свою очередь, вступает в контакт с
ганглиозной клеткой колбочки, что
происходит
в субслое b
внутреннего
плексиформного слоя
[314]. В результате этого формируется так
называемый «возвратный синапс»,
известный
как реципроктный синапс. Эти ганглиоз-ные
клетки отвечают на освещение
деполяризацией,
формируя ON-центр
рецептивного поля. В
то же время амакриновые клетки АН,
контактирующие
с ганглиозной клеткой при помощи
синаптической ленты в субслое а,
отвечают
на
освещение сетчатки гиперполяризацией,
образуя
OFF-центр
рецептивного поля (рис. 4.2.8, 4.2.9). Именно
благодаря этому образуются рецептивные
поля ON/OFF-типа.
Необходимо отметить, что амакриновые клетки АИ также соединяются между собой. Это происходит в субпластинке b при помощи щелевых контактов. Кроме того, амакриновая клетка АН соединяется с биполярной клеткой колбочки. Благодаря приведенной выше схеме нейронных связей амакриновой клетки АН с биполярной клеткой колбочки и ганглиозными клетками сигналы, исходящие от палочек, используются и колбочковой системой при изменении степени освещенности (темновые и мезо-пические условия).
Вторым типом амакриновых клеток являются клетки типа А17. Эти клетки относятся к ГАМК-эргическим нейронам [425, 592] и обладают широким дендритным полем, что позволяет одной клетке объединить до 1000 биполярных клеток палочек. Дендриты амакриновых клеток А17 достигают субпластинки b внутреннего плексиформного слоя, где и завершаются (рис. 4.2.8, 4.2.9). Эти клетки не образуют синапсов с другими амакриновыми и ганглиозными клетками и их функцией является только объединение биполярных клеток палочек, обеспечивая конвергенцию сигнала. По этой причине амакриновая клетка А17 обладает самостоятельным рецептивным полем и регулирует уровень чувствительности палочек и биполярных клеток по площади сетчатки, участвуя тем самым в темновой и световой адаптации. Благодаря конвергенции сигнала эта клетка очень чувствительна к освещению низкой интенсивности.
Третьим типом амакриновых клеток тракта палочек является клетка типа А18, дендритное поле которой распределяется только в субпластинке а внутреннего плексиформного слоя (рис. 4.2.9). Эта амакриновая клетка относится к допаминэргическим (выявляется при проведении иммуногистохимических реакций на ферменты синтеза допамина, тирозин-гидроксила-зы). Отростки клетки типа А18 окружают тела и дендриты амакриновых клеток типа АИ, А8, А17 (рис. 4.2.9). Они также образуют большое количество синапсов с трактами палочек, амакриновыми и биполярными клетками колбочек. Эти амакриновые клетки отвечают на возбуж-
дение деполяризацией. Предполагают, что до-паминэргическая клетка А18 увеличивает размер рецептивного поля, повышая при этом чувствительность ганглиозных клеток в ското-пических условиях. Увеличение рецептивного поля исключительно важно (помимо изменения скорости синтеза пигментов фоторецепторов) в реализации механизмов темновой адаптации.
Система приведенных выше амакриновых клеток обеспечивает не только конвергенцию сигнала, но и его дивергенцию, благодаря обратным связям. Степень конвергенции и дивергенции системы палочек можно проиллюстрировать на примере организации сетчатки кошки [314]. Конвергенция сигнала сводится к тому, что приблизительно 1500 палочек передают информацию отдельной ON-бета ганглиозной клетке посредством 100 биполярных клеток палочек, 5 амакриновых клеток типа АИ и 4 биполярных клеток колбочек. Информация к OFF-альфа ганглиозной клетке поступает от 75 000 палочек, 5000 биполярярных клеток палочек и 250 амакриновых клеток. Дивергенция палочковой системы сводится к тому, что отдельная палочка передает информацию двум биполярным клеткам, а от них 5 амакриновым клеткам типа АИ, 8 биполярным клеткам колбочек и двум ON-бета ганглиозным клеткам. Благодаря существованию дивергентной и конвергентной систем обеспечивается объединение и усиление сигнала палочек при очень низком освещении. Чувствительность столь высока, что зрительная система реагирует на единственный квант световой энергии. Благодаря наличию связи между палочковой и колбочковой системами посредством амакриновых клеток, палочковая нейронная система в мезопи-ческих условиях передает свои функции кол-бочковой системе.
Необходимо подчеркнуть и то, что приведенная выше схема взаимодействия между нейронами палочкового тракта является основой формирования рецептивных полей, обеспечивающих наиболее важные функции зрительной системы — контрастную чувствительность, адаптацию.
Нейронная система колбочек. Нейронная система колбочек обеспечивает наибольшую остроту зрения в фотопических условиях, а также обладает способностью цветовосприятия. В определенных условиях (темновые) колбоч-ковая система регистрирует ахроматические сигналы. Первоначально мы охарактеризуем основные закономерности формирования кол-бочковой системы.
Необходимо отметить, что система прохождения сигналов от колбочки к ганглиозным клеткам довольно существенно отличается от нейронной системы палочек. Так, одна колбочка образует синапсы с двумя биполярными клетками, в то время как палочка только с одной (рис. 4.2.3, 4.2.8). Благодаря этому уже
420