

Глава 4. Головной мозг и глаз
 затора
этот эффект проявляется в виде так
называемого
«последовательного образа». Наиболее
ярким примером существования такого
эффекта
может служить следующее явление. Если
после длительного рассмотрения красного
изображения
перевести взляд на белую поверхность,
белая поверхность будет казаться
зеленой, т. е. цвета, дополнительного
к красному.
Особенно часто приходится сталкиваться
с явлением
последействия движения. Например, при
вращении колеса со спицами с определенной
скоростью возникает впечатление
вращения
колеса в обратном направлении. Явление
последействия
указывает на существование одновременно
двух антагонистических сенсорных
систем,
уравновешенных в обычных условиях.
Можно
предположить, что при длительном
возбуждении
одной из систем световым сигналом ее
чувствительность снижается, а
антогонисти-ческая
система сохраняет свою чувствительность.
Если предположить, что детектор красного
цвета в приведенном выше примере спарен
с детектором зеленого цвета, то оба они
одинаково
реагируют на белый цвет и на выходе
пары нет ничего, кроме фона. Предположим
теперь, что глаз фиксирует красную
поверхность
в течение некоторого времени, в результате
чего рецепторы красного цвета
«утомляются».
Если перевести взгляд на белую
поверхность,
то рецепторы зеленого будут реагировать
нормально, а рецепторы красного дадут
ослабленную реакцию. Таким образом,
свет будет
восприниматься как зеленый. Необходимо
отметить, что приведенное объяснение
легло
в основу оппонентной теории восприятия
цвета,
на чем мы остановимся несколько ниже.
Наличие
детекторов, представляющих собой
организованные
определенным образом рецептивные
поля и обладающих антагонистическими
свойствами, которые реагируют на стимулы
различной
формы (линии, щели, углы и т. п.) и цвет,
было позднее обнаружено нейрофизиологами
на уровне сетчатки, наружных коленчатых
тел и различных участков коры головного
мозга при помощи разработанных методов
электрофизиологии,
позволяющих снимать потенциал
отдельного нейрона. На основе этих
исследований теоретически разработаны
схемы нейронных
цепей, удовлетворяющие требованиям
детекторов различных характеристик
изображения
при его анализе. В последующем
ней-рогистологи при помощи световой,
электронной микроскопии
и методов гистохимии выявили особенности
этих цепей, заключающиеся в определенной
связи между нейронами различных типов,
характере контактов между ними, а также
в использовании нейронами при передаче
нервного
импульса различных нейромедиато-ров.
Основополагающим понятием, возникшим
в
результате приведенных выше наблюдений,
явилось
понятие «рецептивное поле». Ранее мы
неоднократно упоминали термин
«рецептивное
затора
этот эффект проявляется в виде так
называемого
«последовательного образа». Наиболее
ярким примером существования такого
эффекта
может служить следующее явление. Если
после длительного рассмотрения красного
изображения
перевести взляд на белую поверхность,
белая поверхность будет казаться
зеленой, т. е. цвета, дополнительного
к красному.
Особенно часто приходится сталкиваться
с явлением
последействия движения. Например, при
вращении колеса со спицами с определенной
скоростью возникает впечатление
вращения
колеса в обратном направлении. Явление
последействия
указывает на существование одновременно
двух антагонистических сенсорных
систем,
уравновешенных в обычных условиях.
Можно
предположить, что при длительном
возбуждении
одной из систем световым сигналом ее
чувствительность снижается, а
антогонисти-ческая
система сохраняет свою чувствительность.
Если предположить, что детектор красного
цвета в приведенном выше примере спарен
с детектором зеленого цвета, то оба они
одинаково
реагируют на белый цвет и на выходе
пары нет ничего, кроме фона. Предположим
теперь, что глаз фиксирует красную
поверхность
в течение некоторого времени, в результате
чего рецепторы красного цвета
«утомляются».
Если перевести взгляд на белую
поверхность,
то рецепторы зеленого будут реагировать
нормально, а рецепторы красного дадут
ослабленную реакцию. Таким образом,
свет будет
восприниматься как зеленый. Необходимо
отметить, что приведенное объяснение
легло
в основу оппонентной теории восприятия
цвета,
на чем мы остановимся несколько ниже.
Наличие
детекторов, представляющих собой
организованные
определенным образом рецептивные
поля и обладающих антагонистическими
свойствами, которые реагируют на стимулы
различной
формы (линии, щели, углы и т. п.) и цвет,
было позднее обнаружено нейрофизиологами
на уровне сетчатки, наружных коленчатых
тел и различных участков коры головного
мозга при помощи разработанных методов
электрофизиологии,
позволяющих снимать потенциал
отдельного нейрона. На основе этих
исследований теоретически разработаны
схемы нейронных
цепей, удовлетворяющие требованиям
детекторов различных характеристик
изображения
при его анализе. В последующем
ней-рогистологи при помощи световой,
электронной микроскопии
и методов гистохимии выявили особенности
этих цепей, заключающиеся в определенной
связи между нейронами различных типов,
характере контактов между ними, а также
в использовании нейронами при передаче
нервного
импульса различных нейромедиато-ров.
Основополагающим понятием, возникшим
в
результате приведенных выше наблюдений,
явилось
понятие «рецептивное поле». Ранее мы
неоднократно упоминали термин
«рецептивное
поле». Об особенностях функционирования рецептивных полей мы сейчас остановимся более подробно.
Рецептивное поле на уровне сетчатки представляет собой совокупность точек сетчатки (фоторецепторных клеток), в пределах которой зрительный стимул вызывает возбуждение или торможение одного нейрона следующего (более высокого) порядка, в частности ганглиозной клетки.
Еще в 30-е годы прошлого столетия Hart-line [231, 232] развил концепцию рецептивного поля при изучении глаза краба и лягушки. В дальнейшем были выполнены классические эксперименты на кошках [319, 320], сформировавшие базу для последующих исследований зрительной системы млекопитающих.
Как было указано выше, возможность подобных экспериментов появилась в связи с созданием методик подведения микроэлектрода непосредственно к одному нейрону. Именно таким образом регистрируется потенциал действия ганглиозных клеток сетчатки, нейронов наружных коленчатых тел и нейронов зрительной коры головного мозга во время освещения сетчатки паттернами различной формы и плотности энергии (рис. 4.2.5). Это позволило исследователям анализировать как параметры зрительных стимулов, возбуждающих или тормозящих нейроны, так и особенности реакции нейронной системы.
Используя эту методику, первое, что обнаружили Kuffler и Hartline [320], это существование двух категорий ганглиозных клеток. Клетки ON-типа возбуждались при освещении, а OFF-типа возбуждались при отсутствии освещения. Причем участки сетчатки, отвечающие противоположной реакцией, располагались рядом, чаще в виде кольца. Центральная часть такой окружности отвечала на стимул одним типом реакции, а периферия противоположной. При регистрации потенциала действия ганглиозной клетки световое пятно, помещенное
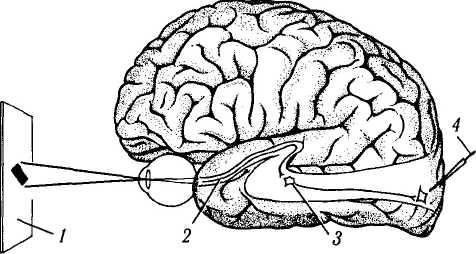
Рис. 4.2.5. Схема проведения электрофизиологических исследований функциональной активности различных отделов зрительного пути (объяснения в тексте):
1 — зрительное поле; 2 — зрительный тракт; 3 — наружное коленчатое тело; 4 — электрод в зрительной коре
Функциональная анатомия зрительной системы
417

в
центр рецептивного поля, вызывает
усиление активности
клетки, т. е. формируется ON-ответ
(рис.
4.2.6, см.
цв. вкл.). Когда
такое же небольшое
световое пятно помещают вне рецептивного
поля, нейрон уменьшат свою активность
(OFF-ответ).
Подобного типа рецептивное
поле называется ON-центр
рецептивным полем.
Существуют и OFF-центр
рецептивные поля,
особенностью которых является торможение
ганглиозной клетки при освещении
центра
рецептивного поля.
Куфлером изучались также рецептивные поля с целью определения «оптимального сигнала», т. е. сигнала в наибольшей степени возбуждающего ганглиозную клетку. Оказалось, что самым сильным сигналом для ON-центр ганглиозной клетки было световое пятно, полностью выполняющее центр рецептивного поля (см. рис. 4.2.6). Таким же образом наиболее оптимальным для тормозного сигнала явилось освещение периферии рецептивного поля стимулом, имеющим вид кольца. После подобного торможения в момент снятия светового сигнала нейрон формировал потенциал действия.
Установлено также, что если освещать рецептивное поле большим световым пятном, накрывающим как центр, так и периферию поля, возникает значительно более слабый ответ, чем при использовании маленького пятна, освещающего только центр. Таким образом, тормозная реакция периферии рецептивного поля ослабляла или даже устраняла центральное возбуждение. Приведенные выше реакции рецептивного поля связаны с особым типом обратных связей тормозного характера между нейронами сетчатой оболочки (рис. 4.2.7).
Из приведенных выше фактов становится ясно, что в основе большинства электрофизиологических особенностей рецептивных полей лежат явления возбуждения и торможения, морфологическим субстратом которых являются обратные связи возбуждающего или тормозного характера между рядом расположенными нейронами. Именно наличием таких связей между нейронами с формированием рецептивных полей можно объяснить феномены, приведенные на рис. 4.2.4.
В настоящее время известно большое количество разнообразных типов реакций зрительной системы, основой которых является функционирование рецептивных полей. Существование рецептивных полей сетчатки определяет фундаментальные функциональные ее свойства, такие как обеспечение одновременного и последовательного зрительного контраста, острота зрения, движение объекта, световая и темно-вая адаптация, обработка информации о цвете объекта. При этом указывают на наличие довольно сложной организации рецептивных полей, выполняющих ту или иную функцию. На уровне сетчатки доказано наличие рецептивных полей, реагирующих на форму объекта (выде-
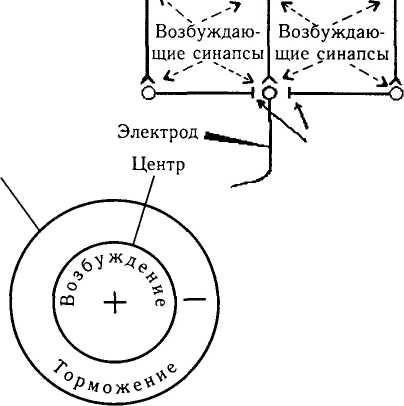
Тормозные синапсы
Окружение
и и
Рис. 4.2.7. Схема обратных свезей между нейронами сетчатки, формирующими ргцептивные поля (объяснение в тексте)
ляя такие признаки, как наличие и длина линий, наличие углов и т. п.), его движение, спектральную характеристику объекта. Благодаря сложному взаимодействию нейронов уже в сетчатке зрительная информация обрабатывается, кодируется и по зрительному нерву передается наружному коленчатому телу [592]. Структура рецептивных полей сохраняется и на уровне наружных коленчатых тел, а также в зрительной коре головного мозга. Именно подобная организация обеспечивает передачу информации по принципу «точка к точке». Этот принцип сводится к тому, что фоторецепторы определенных участков сетчатой оболочки передают информацию ганглиозным клеткам, а затем нейронам наружного коленчатого тела и зрительной коры, сохраняя при этом топографическое пространственное расположение рецептивных полей. Это топографическое распределение не линейное. Примером тому является значительно большее представительство макулярной области в наружном коленчатом теле и коре относительно остальной части сетчатки.
В настоящее время достаточно точно выявлены морфологические основы нескольких нейронных сетей сетчатки, определяющие их различные функции. Это система палочек, система колбочек, «карликовая» система фовео-лярной области, тракт S-колбочек и обратные связи. Особенности организации этих сетей сводятся к наличию определенных типов нейронов, а также синаптических соединений, использующих различные медиаторы.
418