

Глава 4. Головной мозг и глаз
 нение
в связи с тем, что структурные различия
хорошо коррелируют с функциональными
особенностями,
о чем будет изложено несколько
ниже.
нение
в связи с тем, что структурные различия
хорошо коррелируют с функциональными
особенностями,
о чем будет изложено несколько
ниже.
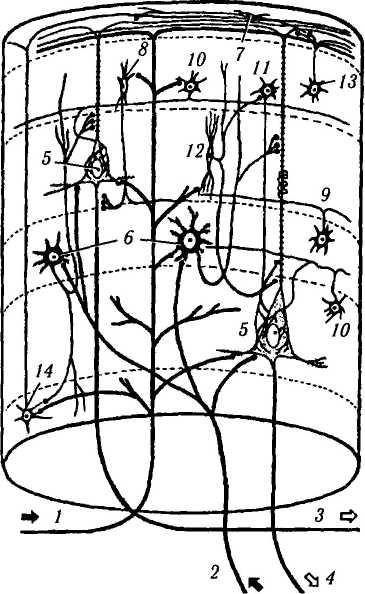
Вертикальная организация коры. Общий поток восходящей и нисходящей информации к коре и от нее организован вертикально. То есть информация, идущая от зрительного бугра, передается звездчатым клеткам IV слоя, которые, в свою очередь, передают ее более высоко и более низко расположенным клеточным слоям для дальнейшей обработки. Подобная вертикальная, или колоночная, организация довольно легко демонстрируется при использовании не только новых морфологических, но и физиологических методов (рис. 4.1.10). Эти методы позволили точно установить, что кору мозга можно подразделить на структурно-функциональные колонки, распространяющиеся от мягкой мозговой оболочки до белого вещества [3, 28, 286]. При этом в границах колонки все нейроны выполняют одну и ту же четко определенную функцию. Ширина такой функциональной ко-
III IV
V VI
Рис. 4.1.10. Колоночная организация коры полушарий большого мозга (по В. J1. Быкову, 1997). На схеме показаны главные ассоциативные связи клеток основных типов:
/ — кортико-кортикальное афферентное волокно; 2 — таламо-кор-тикальное афферентное волокно; 3 — ассоциативное эфферентное волокно; 4 — проекционное эфферентное волокно; 5 — пирамидная клетка; б — шипиковая звездчатая клетка; 7 — горизонтальная клетка Кахала; 8 — аксо-аксонная клетка; 9 — клет-ка-«канделябр», 10—корзинчатая клетка; //—колонковая кор-зинчатая клетка; 12 — клетка с двойным букетом дендритов; 13 — клетка с аксональным пучком; 14 — клетка Мартинотти. Римскими цифрами обозначены слои
лонки колеблется от 250 до 500 мкм, а их количество равняется 2—3 млн. Каждая колонка содержит примерно 5000 нейронов. Внутри колонки выделяют также более мелкие мини-колонки, включающие структуры, непосредственно окружающие апикальные дендриты пирамидных клеток. Колонка включает:
афферентные пути;
систему локальных связей;
эфферентные пути.
Афферентные пути организованы следующим образом. В центре колонки проходит примерно 100 возбуждающих кортико-кортикаль-ных волокон — аксонов пирамидных клеток других колонок данного и противоположного полушарий. Они образуют окончания во всех слоях колонки (в том числе на клетках Мартинотти, шипиковых звездчатых клетках, латеральных дендритах пирамидных клеток) и проходят до I слоя, где образуют ветви, уходящие за ее пределы.
Специфические афферентные импульсы по таламо-кортикальным волокнам поступают на тела и дендриты пирамидных клеток и на шипи-ковые звездчатые клетки IVDcnoH (последние по своим аксонам передают их на апикальные и базальные дендриты пирамидных клеток).
Система локальных связей формируется вставочными нейронами колонки, которые включают более десятка типов клеток. Часть из них обладает тормозной функцией и регулирует преимущественно активность пирамидных клеток. Из тормозных нейронов колонки наибольшее значение имеют следующие [3] (см. рис. 4.1.10):
Аксо-аксонные клетки, тела которых ле жат во II—III слоях, а аксоны идут горизон тально, отдавая многочисленные терминальные веточки, которые образуют тормозные синапсы на начальных сегментах аксонов пирамидных клеток II и III слоев.
Клетки-«канделябры», которые встреча ются во всех внутренних слоях коры. Их аксон- ные коллатерали идут горизонтально и дают несколько восходящих и нисходящих веточек, которые образуют спиральные ветвления во круг апикальных дендритов пирамидных клеток.
Корзинчатые клетки, которые располага ются во II слое, на границе III и IV, а также IV и V слоев. Их аксоны проходят горизонталь но на расстояние до 3 мм и, оплетая тела круп ных и средних пирамидных клеток, влияют на 20—30 соседних колонок. Колонковые корзин чатые клетки обеспечивают торможение пира мидных клеток по вертикали внутри данной колонки.
Клетки с двойным букетом дендритов, от ходящих вертикально от полюсов клетки, рас положены во II—III слоях. Их аксон дает кол латерали, образующие контакты с дендритами как пирамидных клеток, так и непирамидных (в том числе тормозных) нейронов. Первый тип контактов опосредует угнетение пирамидных
Анатомия головного мозга
365
 клеток,
а второй — их активацию путем снятия
торможения.
клеток,
а второй — их активацию путем снятия
торможения.
5. Клетки с аксонным пучком (кисточкой) — звездчатые нейроны II слоя, аксоны которых ветвятся в I слое, образуя связи с дистальными сегментами апикальных дендритов пирамидных клеток и с горизонтальными ветвями кортико-кортикальных волокон.
Эфферентные пути формируются следующим образом. Аксоны средних пирамидных клеток III слоя устанавливают связи преимущественно с соседними колонками и колонками противоположного полушария, а аксоны крупных и гигантских пирамидных клеток V слоя, помимо этого, направляются в подкорковые центры, образуя вместе с аксонами веретено-видных клеток VI слоя систему эфферентных волокон коры.
Определенные особенности вертикальной организации коры существуют в области зрительной коры. Эти сведения будут приведены в разделе, посвященном зрительной коре.
Приводящие и отводящие связи коры. Нейроны коры можно подразделить на ряд категорий в соответствии с особенностями их связей. К первой категории нейронов относятся проекционные нейроны, которые передают импульсы к подкорковым центрам, таким как зрительный бугор, ствол мозга, спинной мозг или базальные ганглии. Ко второй категории относятся вставочные нейроны, формирующие связи с другими нейронами в том же самом полушарии. К третьей категории относятся ко-миссурные нейроны, которые напоминают по
функции вставочные нейроны за исключением того, что они посылают аксоны в противоположное полушарие через мозолистое тело или переднюю спайку мозга [3, 4, 6, 8].
Большинство афферентов заканчивается в
IV слое коры головного мозга (рис. 4.1.11). Эфференты, исходящие из коры, начинаются от нейронов V и VI слоев. Пирамидные нейроны
V слоя проецируются непосредственно на ствол мозга или спинной мозг. Нейроны VI слоя явля ются основным источником прямых проекций на зрительный бугор. Эфференты, направляю щиеся к другим областям коры (т. е. вставоч ные нейроны), исходят от нейронов III и VI слоев.
Архикортекс. Филогенетически наиболее старые структуры коры названы палеокортек-сом, мезокортексом и архикортексом. Они характеризуются слоистой организацией, но число слоев нейронов в этих участках коры меньше шести. Основной областью палеокортекса является обонятельная кора, мезокортекса — кора поясной извилины, а основной частью архикортекса — гиппокамп. Гиппокамп отличается от коры обратным расположением слоев. При этом серое вещество гиппокампа располагается внутри, а белое — снаружи. Часть гиппокампа состоит только из трех слоев клеток.
Нейромедиаторная система коры. В коре выявлено более десятка различных нейроме-диаторов и рецепторов к ним. Это норадрена-лин, серотонин, ацетилхолин, вазоинтерстици-альный кишечный полипептид (VIP), соматоста-тин, холецистокинин, глютамат и др.
Слои
Афференты
Эфференты
Мягкая оболочка
 III
III
А
А
Кора мозга
IV

 VI
VI
А
 Белое
вещество
Белое
вещество



 V
V
V
V
Кортикокор-тикальные
Кортикокор-тикальные
Таламокор-тикальные
Кортикотала- Кортикобульбарные мические Кортикопонтинные
Кортикоретикулярные
Кортикоспинальные
Кортикостриарные
Рис. 4.1.11. Особенности связей коры головного мозга (объяснение в тексте)
366