

Глава 4. Головной мозг и глаз
 является
сопоставление сведении, получаемых
при анализе результатов аутопсии, с
результатами
экспериментальных исследований.
является
сопоставление сведении, получаемых
при анализе результатов аутопсии, с
результатами
экспериментальных исследований.
Каким же образом происходит формирование зрительного пути человека? Ганглиозные клетки сетчатки человека возникают между восьмой и пятнадцатой неделями беременности. Наибольшее количество ганглиозных клеток (2,2 до 2,5 млн) обнаруживается на 18-й неделе развития и остается таковым до 30-й недели. Затем их количество резко снижается в результате гибели части нейронов. В последующем количество клеток также постепенно, но медленно снижается. Продолжается процесс уменьшения числа клеток и после рождения на протяжении нескольких месяцев. В результате этого число ганглиозных клеток сокращается до 1 млн. Объясняют гибель ганглиозных клеток необходимостью «улучшения» формирования проекций сетчатой оболочки на наружное коленчатое тело, что, возможно, связано с удалением несущественных связей.
В данном месте необходимо остановиться на особенностях развития нейронов зрительного пути обезьяны, основного объекта экспериментальных исследований. Сравнивая этапы развития той или иной структуры у обезьяны, возможно экстраполировать, с определенной осторожностью, получаемые результаты и на человека.
У обезьян нейроны наружного коленчатого ядра начинают формироваться между 36-м и 43-м днями, а беременность продолжается только 165 дней. Учитывая разность в длительности беременности у обезьян и человека, можно считать, что 36—43-е сутки у обезьяны соответствуют 8—11-й неделям эмбриогенеза человека. Таким образом, наружное коленчатое тело у человека начинает формироваться между 8—11-й неделями эмбрионального развития. Уже к 10-й неделе первые аксоны ганглиозных клеток сетчатки проникают в наружное коленчатое тело.
Сегрегация аксонов ганглиозных клеток сетчатки происходит параллельно с формированием слоистости наружного коленчатого тела. У зародыша человека пластины наружного коленчатого тела и сегрегация афферентов появляются между 22-й и 25-й неделями.
«Подключение» нейронов к первичной зрительной коре возникает между 43-ми и 102-ми сутками (обезьяна). Этот период у человека соответствует от 10 до 25 неделям эмбрионального развития.
Инъекцирование радиоактивных трейсеров показало, что первоначально афференты наружного коленчатого тела, представляющие каждый глаз, интенсивно распространяются в IVC-слое коры. Формирование «глазодоминант-ных колонок» выявляется в течение последних нескольких недель беременности и почти завершается к моменту рождения [333]. Развитие
«глазных доминантных колонок» трудно исследовать у обезьян и значительно легче у кошек (у кошек после рождения происходит формирование НКТ и зрительной коры, что позволяет проводить эксперименты постнатально). По этой причине подобные исследования проведены на котятах.
Внутриглазная инъекция Н-пирролидин-аль-фа-карбоновой кислоты в одно из глазных яблок при 15-дневном возрасте котенка (спустя неделю после открытия у него глаз) приводит к накоплению изотопа в IV слое (рис. 4.2.64). В последующие несколько месяцев постепенно начинают появляться в этом слое «глазодоми-нантные колонки». Светлые полосы выявляют афференты наружного коленчатого тела, соответствующие тому глазному яблоку, в которое

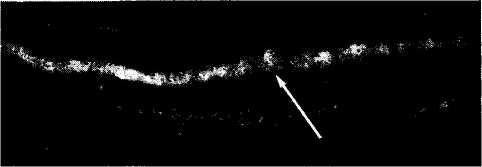
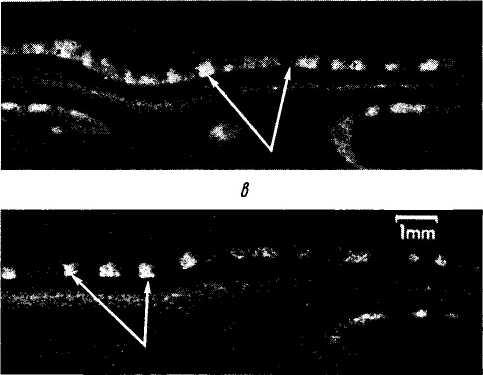
Рис. 4.2.64. Постнатальное развитие «глазодоминант-
ных колонок» кошки (в одно глазное яблоко введен
радиоактивный пролин) (по LeVay, Stryker, Shatz,
1978):
а— 15-й день; б — 22-й день; в — 39-й день; г — 92-й день. К 15-му дню афференты распространяются вдоль IV слоя. В последующем происходит прогрессивная агрегация афферентов с формированием доминантных «колонок», которые становятся все более четкими в виде белых полосок (стрелки)
Функциональная анатомия зрительной системы
463

 был
введен изотоп. Темные полосы принадлежат
второму глазу.
был
введен изотоп. Темные полосы принадлежат
второму глазу.
Созревание глазных «доминантных колонок» сопровождается формированием многочисленных терминалов в IV слое первичной зрительной коры. У 17-дневного котенка геникулокор-тикальные волокна разветвляются только в однородной области IV слоя.
Необходимо отметить, что в первые недели после рождения «глаздоминантные колонки» не совсем четкие [333]. Четкая кластеризация колонок обнаруживается только у взрослых животных. Показано, что формирование «глазо-доминантных колонок» происходит параллельно с дифференциацией дендритического поля нервных волокон наружного коленчатого тела.
Вышеприведенные особенности структурного развития зрительного пути предполагают наличие низких зрительных функций на момент рождения. Каждому офтальмологу известно, что зрительные функции у новорожденных довольно примитивны (низкая острота зрения). Установлено, что острота зрения у новорожденных всего 5% и довольно быстро увеличивается до 100% на протяжении нескольких первых лет жизни. Это быстрое развитие зрительных функций происходит параллельно «созреванию» механизмов, управляющих аккомодацией, бинокулярным зрением, движениями глаза [64, 167].
Выявлено, что совершенствование зрительных функций после рождения сопровождается анатомическими изменениями, которые происходят одновременно на всех уровнях зрительного пути — от сетчатой оболочки до зрительной коры. Так, у человека при рождении ма-кула не полностью сформирована. Имеет она многослойное строение, а число колбочек незначительно. В течение первого года жизни фоторецепторы перераспределяются в пределах сетчатки, а плотность колбочек в фовеа увеличивается в пять раз. У новорожденных аксоны плохо миелинизированы, но уже в течение первых двух лет этот процесс проходит быстрее и завершается на первом десятилетии жизни.
Наружное коленчатое тело при рождении содержит только около 60% нейронов, и их количество увеличивается на протяжении 2 лет.
Плотность дендритных полей и синапсов достигает пика только в 8-месячном возрасте, а совершенствование синаптических связей первичной зрительной коры происходит на протяжении многих лет после рождения.
Для полноценного структурно-функционального становления зрительного пути необходима «полноценная» зрительная стимуляция системы. Показано, что зрительная система начинает формироваться у эмбриона раньше, чем могут оказать влияние на этот процесс «зрительные стимулы». Современные данные позволяют предположить, что после рождения существуют механизмы, участвующие в процессах
«созревания» зрительной системы. Для выявления этих механизмов Wiesel и Hubel поставили эксперименты, которые сводились к тому, что новорожденным обезьянам зашивались веки. Они установили, что у новорожденных можно обнаружить «простые» и «комплексные» клетки, напоминающие аналогичные у взрослых животных. В первичной зрительной коре определяются как «ориентационные колонки», так и «глаздоминантные колонки», которые фактически не отличались от таковых у взрослых обезьян. Полученые данные позволили заключить, что основные структурно-функциональные элементы зрительной коры уже сформированы перед рождением благодаря генетически детерминированным командам.
Странным оказалось то, что физиологическая активность зародыша играет большую роль в развитии нормальных анатомических связей в зрительной системе. Внутриутробно ганглиоз-ные клетки сетчатки самопроизвольно формируют потенциалы действия в отсутствии какой-либо зрительной стимуляции. Отменяя возможность формирования этих потенциалов тетро-доксином (блокирует натриевый канал), можно добиться нарушения нормальной пренатальной сегрегации аксонов ганглиозных клеток сетчатки, формирования пластин наружного коленчатого тела и «глазодоминантных колонок». Эти эксперименты указывают, что, хотя функциональная архитектура зрительной системы детерминирована генетически, «усовершенствование» связей обеспечивается физиологической активностью, существующей уже у зародыша.
Критический период. Если новорожденную обезьяну содержать в темноте или на оба века наложить швы, формирующиеся рецептивные поля первичной зрительной коры имеют довольно причудливые свойства. Исчезают их ориентационные свойства и нормальные бинокулярные ответы. Часть клеток вообще не отвечает на стимуляцию зрительным объектом, и они могут быть обнаружены только на основании их аномальной самопроизвольной активности. Если после такой длительной депривации обезьяну поместить в нормальные условия зрительной активности, нейроны первичной зрительной коры не восстанавливают нормальную реакцию на зрительные объекты.
Эти исследования показывают, что для формирования нормальной зрительной функции требуется стимуляция системы специфическими раздражителями на протяжении определенного (критического) периода после рождения.
У человека определен критический период на основании анализа результатов хирургического удаления врожденных катаракт, выполненного в различном возрасте. Критический период у человека колеблется в довольно широких пределах и может составлять несколько лет после рождения [220, 332, 497].
•им