Запитання для самоперевірки
Що вивчає фізіологія риб?
Які основні завдання стоять перед фізіологією риб?
Яка основна мета вивчення фізіології риб?
Які основні підходи до вивчення функцій риб та які розділи фізіології їх вивчають?
Опишіть основні методи дослідження в фізіології.
Які основні особливості проведення досліджень на рибах?
Які основні особливості водного середовища як середи існування?
Які особливості складу тіла риб?
Чим відрізняються живі організми від неживих? Перелічити основні властивості живого організму.
Що таке гомеостаз?
Як здійснюється саморегуляція функцій?
Які основні принципи регуляції процесів в організмі риб?
Яка різниця в поняттях “гуморальна” і “гормональна” регуляція?
Що таке негативний зворотний зв’язок?
Що таке функціональна система?
Розділ 2. Фізіологія збудження Основні функції клітини
Клітина є основою будови і функції живого організму, вона лежить в основі всіх складних комплексів (функціональна одиниця, система органів, функціональна система), які забезпечують його життєдіяльність.
Стабільна система роботи клітини забезпечується клітинними системами саморегулювання, але поряд із цим зміна функціонального стану клітини забезпечується за рахунок гуморальної і нервової регуляції, які можуть змінювати її функціональний стан залежно від фізіологічного стану організму.
Зв’язок клітини із іншими клітинами, їх реагування на зміни у міжклітинному середовищі і регуляція забезпечується структурами, що розташовані в мембрані клітини.
Мембранні структури
Мембрани є основним структурним елементом клітини. Плазматична мембрана відокремлює клітину, її цитоплазму від навколишнього середовища. Усі клітинні органели – ядро, ендоплазматичний ретикулум, апарат Гольджі, мітохондрії, пластиди, мікросоми, лізосоми тощо побудовані із мембранних структур. Мембрани являють собою складну систему, наділену ферментативною активністю, яка здійснює координований поліферментний каталіз. При всій розмаїтості устрою мембран і їхньої біологічної ролі усі вони улаштовані за єдиним принципом і займають від 1/3 до 1/2 маси клітини.
Мембранні структури забезпечують розмежування внутрішньоклітинного простору на особливі відсіки – компартменти, усередині кожного із який здійснюється свій визначений цикл реакцій, а взаємодія між ними реалізується через регульовану перемінну проникність відповідних мембран.
Основою біологічних мембран усіх типів є ліпідний шар. Саме ліпіди легко утворять плівки на границі двох середовищ, двох розчинів, і еволюційно, імовірно, із утворення таких ліпідних поверхонь роздягнула і почалося утворення протомембран і протоклітин. До складу мембран обов'язково входять фосфоліпіди, холестерин, ліпопротеїди і білки, кількість яких варіює. Ліпіди складають 40% сухої маси мембран, причому 80% із них приходиться на частку фосфоліпідів. Саме фосфоліпіди утворюють подвійний молекулярний шар, що відокремлює компартменти клітини друг від друга і клітинну цитоплазму від позаклітинного середовища. Фосфоліпіди в бімолекулярному шарі розташовані паралельно один одному й утворять дві гідрофобні (водовідштовхувальні) поверхні, які пролежать один до одного. Гідрофільні ж, полярні групи ліпідів, звернені назовні бімолекулярного шару. Таким чином, подвійний гідрофобний шар мембрани є ізолюючим прошарком, що перешкоджає пасивному транспорту води, електролітів, кисню і метаболітів (за винятком жиророзчинних).
Зовсім необхідне для нормального функціонування мембрани властивість виборчої проникності реалізується за рахунок існування в структурі мембран проміжків в упакуванні молекул фосфоліпідів, появи полярних угруповань у гідрофобному шарі (пасивний транспорт), а також функцією спеціалізованих білків-каналоутворювачів, що здійснюють активний транспорт іонів і метаболітів, у тому числі і проти градієнта концентрацій.
Щільність упакування подвійного ліпідного шару мембран залежить від довжини ланцюга молекул (чим ланцюги довше, тим щільніше упакування). Присутність ненасичених жирних кислот і відповідних фосфоліпідів робить подвійний ліпідний шар більш розтягнутим, менш щільним. Щільність упакування ліпідних молекул залежить і від кількості присутнього в мембранах холестерину.
При біологічних температурах мембранні ліпіди, принаймні в гідрофобному центрі подвійного ліпідного шару, знаходяться в рідкокристалічному стані, а при зниженні температури переходять у кристалічний стан. Двовалентні катіони Са2+ і Mg2+ підвищують температуру переходу, нейтралізуючи негативний заряд "голівок" фосфоліпідів. Одновалентні катіони Li+, Na+, К+ знижують температуру переходу. Плинність – латеральна рухливість мембранних ліпідів – відіграє важливу роль у реалізації функцій мембран, зокрема, транспортної і рецепторний. Рухливість іонофорів (каналоутворювачів) знеобхідна для їхнього функціонування. Плинність мембран обумовлена, в основному, концентрацією присутнього в них холестерину. Чим вище концентрація холестерину, тим нижче температура фазового переходу.
Природні фосфоліпіди в мембранах при звичайній температурі і надлишку води знаходяться в рідкокристалічному стані, при якому окремі молекули ліпідів (і білків) можуть переміщатися уздовж подвійного шару (латеральна дифузія). Під час зниження температури досягається фазовий перехід ліпідів у твердий желеподібний стан.
Температура фазового переходу залежить від ступеня гідрофобності фосфоліпідів і форми їхніх полярних "голівок". Присутність подвійних зв'язків, укорочення і розгалуження вуглеводневих ланцюгів жирних кислот знижує температуру фазового переходу, послабляє сили вандерваальсових взаємодій між молекулами і збільшує їхню рухливість.
Якщо температура вища за температуру фазового переходу, ліпідна фаза мембран знаходиться в рідинному стані, мірою якої є плинність – здатність пластично чи грузько формуватися під впливом механічної напруги. Плинність обернено пропорційна в'язкості. Величина плинності біомембрани, її подвійного ліпідного шару пропорційна ступені ненасиченості жирнокислотних ланцюгів фосфоліпідів, які до неї входять. Збільшення змісту сфінгомієлину і холестерину знижує плинність мембран. Одночасно зростає відносна непроникність мембран.
У риб, які є пойкилотермними тваринами, плинність мембран змінюється за рахунок включення в їх структуру різних ліпідів, тому плинність їх мембран під час зміни температури навколишнього середовища залишається на постійному рівні, але різке зниження температури може привести до загибелі риби через різке зниження плинністі.
Розташування ліпідів у подвійному шарі може бути асиметричним. На зовнішній стороні подвійного ліпідного шару звичайно переважають холинутримуючі фосфоліпіди, тоді як фосфатидилетаноламіни і серинінозитоли зв'язані більше із внутрішньою поверхнею. Тому зовнішня поверхня подвійного шару мембрани має меншу плинність, що може відігравати визначену роль у виконанні захисної, бар'єрної функції мембрани.
Подвійний ліпідний шар мембран, насамперед плазматичних, виконуючи разом із мембранними білками барьєрно-транспортну функцію, є основою для створення трансмембранної різниці електричних потенціалів (20 мВ) і високого мембранного опору. Нарешті, ліпідна фаза мембран виконує регуляторну функцію. Її рідкокристалічна структура дозволяє швидко й обернено змінювати функції ліпідів, конформацію і властивості мембранних білків, інших компонентів клітинної поверхні. Ліпідний склад мембран також може змінюватися, але більш повільно, чим синхронні зміни плинності, викликані, наприклад збільшенням концентрації Са2+. Постійне відновлення ліпідного складу мембран зв'язано із функцією фосфоліпаз (насамперед фосфоліпази А2). Ця функціональна система здатна адаптивне змінювати склад ліпідів мембран із урахуванням зовнішніх впливів. Так, за зниження температури середовища в ліпідах зростає концентрація поліненасичених жирних кислот, що знижують температуру фазового переходу і сприяють збереженню плинності мембрани. Крім фосфоліпідів у біомембранах присутні ліпіди й іншіх класів – похідні сфінгозина, гліколіпіди, стерини, насамперед холестерин.
Товщина біологічних мембран коливається в межах 4.0 – 13.0 нм, вона залежить, насамперед, від щільності упакування ліпідних молекул.
Поряд із латеральною дифузією в мембранах можливо також вертикальне переміщення молекул ліпідів ("фліп-флоп") – перехід із однієї сторони подвійного ліпідного шару на іншу із частотою один раз за годину чи рідше – за 5 – 6 годин. Менш важлива коливальна рухливість молекул і їхнє обертання. Рухливість молекул – необхідна умова стабільної структури мембран. Наявність варіацій ліпідного і білкового складу різних мембран і навіть різних ділянок однієї мембрани є причиною того, що в однакових фізіологічних умовах різні області мембрани можуть відрізнятися по своїй латеральній рухливості, ступеню структурованості, а отже, і функціональним можливостям. Полярні "голівки" молекул фосфоліпідів несуть електричні заряди, переважно негативні, що створює різницю потенціалів на поверхні розділу і змінює в такий спосіб концентрацію електролітів у поверхні розділу фаз, що приводить до зміни фізичних властивостей середовища, що в остаточному підсумку впливає на функції мембран.
Постійними компонентами структури мембрани є вуглеводи. присутні в ній, головним чином, у виді глікопротеїнів.
Практично всі мембрани на 95% складаються із ліпідів і білків, причому більш половини маси мембрани складають білки. Більшість мембранних білків, поряд зі структурною функцією, грають активну специфічну роль
Вони служать каталізаторами хімічних реакцій, що протікають у клітці, (ферментна функція), взаємодіють із антигенами, гормонами, іншими біологічно активними речовинами (рецепторна функція), виконують функцію елементів, що довідаються, у мембранному транспорті, піноцитозі і хемотаксисі, здійснюють трансмембранний перенос електролітів і інших низькомолекулярних речовин, міжклітинні взаємодії й ін.
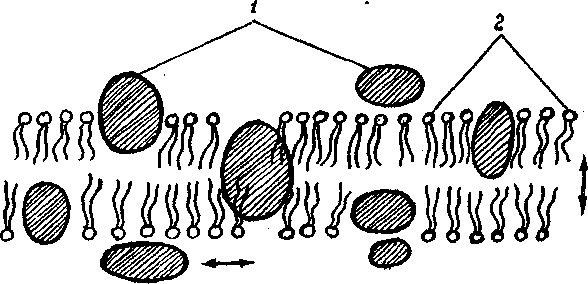
Білкові молекули розташовуються, головним чином, в області полярних "голівок" подвійного фосфоліпідного шару із орієнтацією зовні мембрани. Відповідно білкові молекули можуть брати участь у процесах, що протікають по обох сторони біомембрани. Ступінь зв'язку білка із мембраною може бути різної. Багато мембранних білків своєю гідрофобною частиною закріплені в гідрофобному ліпідному шарі, а їх гідрофільні активні центри звернені назовні мембрани і можуть утворювати навіть активні виступи, мікроворсинки, що забезпечують оптимальні умови для реалізації ферментної, рецепторної і т.п. функцій.
Відомі кілька десятків білків, що входять до складу різних мембран і які виконують різні функції. За характером структурної взаємодії із подвійнім ліпідним шаром розрізняють інтегральні білки (на їхню частку приходиться близько 10% поверхні клітини), що міцно взаємодіють із ліпідними молекулами і які перетинають мембрану по всій її товщині один чи кілька разів; і периферичні білки ( що складають 47% поверхні), зв'язані лише із однієї із поверхонь подвійного шару (мал 2.1). Ці останні легко втрачають зв'язок із мембраною вже при слабких впливах – осмотичному шоку, зміні кислотності середовища, концентрації Са2+ і т.п. Інтегральні білки виділяються із трудом, і при цьому порушується структура мембрани. Склад білків визначає функціональну активність мембран.