

Основні властивості збудливих тканин
Основною властивістю живих систем є здатність відповідати на вплив навколишнього середовища активною реакцією. Особливо яскраво ця властивість виявляється в реакціях нервової, м'язової і залозистої тканин, які називають збудливими, тобто здатними відповідати збудженням на вплив подразників. Подразнення зумовлює в клітині складний комплекс мікроструктурних перебудов, а також зміни обміну речовин, концентрації й швидкості рухові іонів та їх розподілу на клітинних мембранах.
Жива клітина при здійсненні своїх функцій безперервно генерує і підтримує електричні потенціали, тому універсальним способом з'ясування механізмів діяльності збудливих структур є електрофізіологічні дослідження. Основними йонами, які беруть доля в генерації електричних потенціалів клітин, є К+, Nа+ , Са+ і Сl–. За умовами виникнення в живих структурах розрізняють мембранні потенціали спокою (МПС) та потенціали дії (ПД).
Мембранний потенціал спокою
Мембрана клітини електричне поляризована, і різницю потенціалів між зовнішньою і внутрішньою пластинками (поверхнями) клітинної мембрани називають мембранним потенціалом спокою (МПС). Для того щоб зрозуміти природу виникнення цього потенціалу, треба розглянути деякі властивості самої мембрани, а також склад поза- і внутрішньоклітинної рідин.
Клітинна мембрана має значний опір і ємність. Так, у аксона кальмара питомий опір клітинної мембрани (Rм) становить у стані спокою 1000 Ом • см2, а ємність (См) — приблизно 1 мкФ/см2. Ємність мембрани, яка створюється переважно її ліпідним матриксом, є досить сталою, а опір значною мірою залежить від стану її іонних каналів (мал 2.2).
Існування МПС зумовлене передусім неравномірною концентрацією іонів у поза- та внутрішньоклітинному просторі.
Другим важливим чинником, який має значення для створення МПС, є різна проникливість клітинної мембрани для різних іонів. Клітинна мембрана легко проникна для жиророзчинних речовин, молекули яких проходять крізь її ліпідний матрикс. Водорозчинні молекули великого діаметра, у тому числі й аніони органічних кислот, практично не проникають крізь мембрану У стані фізіологічного спокою клітинна мембрана є проникною для К+, оскільки в ній є спеціальні калієві канали, що пропускають лише гідратований калій.
Коли К+ виходить із клітини, він затримується на зовнішній поверхні клітинної мембрани, створюючи її позитивний заряд. Великі аніони не можуть пройти із клітини скрізь мембрану і накопичуються біля її внутрішньої поверхні, створюючи негативний потенціал. Так виникає трансмембранна різниця потенціалів, або МПС.
Цей потенціал протидіє подальшому виходу К+ за електрохімічним градієнтом.
Переміщення К+ із клітини назовні зумовлюється його концентраційним градієнтом, який здійснює осмотичну роботу.
Унаслідок МПС К+ частково повертається назад у клітину. При цьому здійснюється електрична робота.
.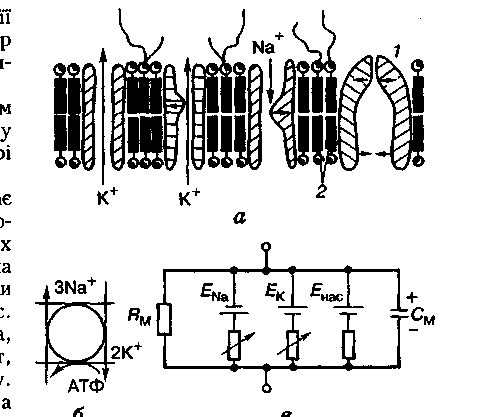
Мал. 2.2. Будова клітинної мембрани (о), її еквівалентна електрична схема (е) і схема активного перенесення йонів за рахунок енергії АТФ (б): / — іонний насос; 2 — ліпідний матрикс (білкові молекули, то утворюють іонні канали і насос, заштриховані); горизонтальними стрілками показано ворітні механізми, вертикальними — напрямок руху іонів; ЕNа, ЕК, Енас — потенціал натрію, калію, насоса; Rм, См — опір та ємність клітинної мембрани
Якщо вихід К+ із клітини переважає над надходженням, то поступово зростає Аелі дещо зменшується аосм. Внаслідок цього на клітинній мембрані виникає потенціал Е, за якого Аел=аосм для К+ і який називають калієвим рівноважним потенціалом (ЕК).
Різницю між поточним значенням МПС і Ек називають електрохімічним градієнтом для КК. Він є основною причиною пасивного руху К+ крізь мембрану в природних умовах.
Якщо відношення внутрішньої і зовнішньої концентрації йонів калію становить, наприклад, 39, а температура середовища + 20˚ С то Ек становитиме 0,092 В.
За такого значення Ек різниця концентрацій К+ між внутрішнім і зовнішнім середовищами клітини залишається практично незмінною, незважаючи на високу проникливість клітинної мембрани до К+, оскільки підтримується постійною роботою натрій-калієвого насоса.
На підтримання МПС витрачається дуже мало К+, основна їх частина повинна забезпечувати електронейтральність внутрішнього середовища клітини. Адже внутрішньоклітинні аніони – це переважно великі білкові молекули, які не можуть пройти крізь клітинну мембрану, і тому їх концентрація залишається сталою. Ці великі аніони мають бути зрівноважені відповідною кількістю катіонів. У зв'язку із тим, що вміст Nа+ всередині клітини дуже незначний, цю функцію виконують К+. Тому внутрішньоклітинна концентрація К+ має бути такою ж високою, як і концентрація великих аніонів, і такою ж стабільною.
МПС створює електричне поле значної напруги — 105 В/см2. Це поле діє на макромолекули мембрани й надає їхнім зарядженим групам певної просторової орієнтації. Крім того, електричне поле МПС забезпечує функцію іонних каналів клітинної мембран.
У стані спокою клітинна мембрана є добре проникною не лише для К+, вона :певною мірою проникна також для інших іонів, зокрема Nа+, С1–, Са2+. Потоки цих іонів порушують рівноважний стан. Той факт, що в умовах спокою Nа+ постійні надходить у клітину, а К+ виходить із неї має дуже важливі наслідки, оскільки система не може бути фактично врівноважена, за допомогою звичайної дифузії і простого відновлення заряду на клітинній мембрані. Якщо немає інших процесів, то клітина дуже швидко накопичуватиме Nа+ і втрачатиме К+, що зменшить їх трансмембранний градієнт. Отже, цей процес може призвести до різкого зниження МПС і загибелі клітини. Проте такого не відбувається, оскільки цьому протидіє натрій-калієвий насос.
Оскільки Nа+ сам не може вийти із клітини, а К+ — надійти в клітину проти своїх електрохімічних градієнтів, вони повинні переміщуватися крізь мембрану активно і на забезпечення цього транспорту має витрачатися енергія. Такий механізм дістав назву натрій-калієвого насоса. Він працює, споживаючи енергію АТФ, і його основним компонентом є фермент мембранна натрій-калій-АТФ-аза. Для роботи насоса обов'язково мають бути зовні іони К+, а всередині — Nа+.
Енергія розщеплення однієї молекулі АТФ забезпечує виведення із клітині трьох іонів натрію і введення всередині клітини двох іонів калію. Це означає, що натрій-калієвий насос є електрогенним: він створює на клітинній мембрані додаткову різницю потенціалів Енас, яка додається до рівня МПС.
Цей електрогенний додаток до нормального рівня МПС неоднаковий для різних клітин. Зазвичай у нервових клітин хребетних він незначний, а в гладком'язових клітинах може становити до 25 % значенні МПС.
Отже, у формувати МПС натрій-каліевий насос виконує дві функції: 1) підтримує трансмембранний градієнт концентрацій Nа+ і К+; 2) генерує певну різницю потенціалів, яка додається до потенціалу що створюється дифузією К+ за концентраційним градієнтом.
Важливим показником здатності іонів проходити крізь клітинну мембрану є її проникність. Значення проникності для іонів К+,Nа+ і Сl– (РК РNа, РСl) можна використовувати для обчислення МПС мембрани, проникної для всіх цих іонів, за рівнянням Гольдмана — Ходжкіна— Катца.
Мембранним потенціалом спокою (МПС) для різних тканин складає:. 20 мВ в епітеліальних клітинах, 60 – 70 в нервових та 80 – 90 в м’язових.