

Хеморецепторні сенсорні системи
До хеморецепторних органів риб відносяться нюх, смак і так зване загальне хімічне відчуття. Звичайно прийнято вважати, що нюх є дистантним органом сприйняття, а смак – контактним. При аналізі реакцій риб на хімічні подразники практично дуже важко, а часом просто і неможливо диференціювати роботу загально хімічних і смакових органів.
Смак
Смакові цибулини локалізовані головним чином у порожнині рота й у районі ротового отвору. Вони входять до складу спеціалізованих хеморецепторних структур – смакових цибулин. Це епітеліальні утворення округлої чи колбоподібної форми, розташовані в товщі багатошарового епітелію.
Смакові цибулини орієнтовані перпендикулярно шарам епітелію так, що апікальний відділ кожної цибулини стикається із навколишнім середовищем. Базальна частина цибулини граничить зі сполучною тканиною, відділеною базальною мембраною. До складу цибулини входить кілька типів клітин: рецепторні вторинночутливі клітини, опорні, базальні і перигамальні. Кожна цибулина включає кілька десятків клітин. Рецепторні клітини мають численні синаптичні контакти із закінченнями аферентних і еферентних волокон.
В еволюційному ряді хребетних смакова рецепція добре розвинена у риб. У водному середовищі роль смакової системи не обмежується харчовою поведінкою. Вона має визначене значення для орієнтації в ближньому полі, оцінки придатності води для дихання, наявності в ній шкідливих сполук і т. ін.
Кількість і локалізація смакових цибулин корелює із екологічними особливостями риб і способом харчування. У придонних риб – мешканців каламутних вод, що харчуються малорухомими тваринами і рослинами, – смакові цибулини крім ротової порожнини, глотки і переднього відділу стравоходу розташовуються по всій поверхні тіла, а також на зябрах і плавцях. У хижих риб смакові цибулини розташовуються лише в порожнині рота і передньому відділі глотки. Смакові цибулини в риб включають кілька десятків рецепторних клітин, що мають на апікальному кінці ряд цитоплазматичних відростків довжиною близько 5 мкм. Всередині відростків розташований пучок паралельно орієнтованих фібрил. У цій області здійснюються первинні процеси хеморецепції.
Смакові цибулини, які розташовані в ротовій порожнині, у глотці й у піднебінному органі, іннервуються 9 і 10 парами черепно-мозкових нервів, а смакові цибулини поверхні тіла і вусиків – 7 парою. Центральні відростки цих нервів закінчуються у відповідних долях довгастого мозку. Лицьові і вагусні долі в риб є первинними смаковими центрами. У них відбувається синаптичне переключення на нейрони вищого порядку. Адекватне подразнення смакових рецепторів викликає зміну електричної активності клітин покришки переднього і середнього мозку. Смакова система риб має представництво в основних інтегративних центрах, що забезпечує її участь у формуванні пристосувальної поведінки.
За допомогою органів смаку риба спроможна уловлювати зміни концентрації солоності води до 0,3 %, вуглекислоти – до 0,8 мг/л, кислотності – до 0,05 одиниці рН. Вона чітко розділяє чотири основні смакові якості: солодке, кисле, солоне і гірке. При цьому чутливість до солодкого, наприклад цукру, у неї майже в 500 разів вище, ніж у людини. Деякі риби дуже тонко відчувають присутність у воді алкоголю. А у окремих випадках риби можуть виявити хімічні об'єкти, особливо харчового характеру, не тільки при контакті, але і на відстані, тобто дистантно.
Нюх
Відповідно до функціональних особливостей органа нюху риб розділяють на макро- і мікросматиків. Нюх перших характеризується найбільш високою чутливістю. Поріг сприйняття цих риб 10 -10 мг/л. Іноді в літературі для образної демонстрації високої чутливості риб цілком виправдано відзначають, якщо в велике озеро під одним берегом випустити одну – дві краплі якоїсь речовини, то риба виявить її присутність під протилежним берегом. Чутливість риб до розчинів чистих хімічних сполук звичайно нижча, ніж до складного біологічно значного запаху. Відмінна риса нюху риб – повільна адаптація.
Запропонована ще більш детальна класифікація, за котрою риби поділяють на 4 групи: гіперосматиків, макросматиків, медіосматиків і мікросматиків. До групи гіперосматиків віднесені представники хрящових риб (акули, скати) із площею нюхового епітелію 2000 – 4350 мм2. Для них характерні домінування нюху, гіпертрофія розеток, цибулин і часткою кінцевого мозку. До групи макросматиків відносяться костисті риби, у поведінці яких нюх має головне значення: площа нюхового епітелію 180 – 430 мм2. Це стайні й одиночні донні і придонні бентофаги – липарис, тріска й ін. У групу медіосматиків виділяються групи риб, що характеризуються помірними параметрами нюхових аналізаторів, площа нюхового епітелію 100 – 265 мм2.
Таблиця 4.1 Площа нюхового епітелію різних груп риб
|
Групи |
Площа нюхового епітелію, мм2 |
Вид риби |
|
Гіперосматики |
2000 – 4350 |
хрящові риби (акули, скати) |
|
Макросматики |
180 – 430 |
костисті (липарис, тріска) |
|
Медіосматики |
100 – 265 |
оселедець, минтай |
|
Мікросматики |
5 – 80 |
стихей, люмпен |
Х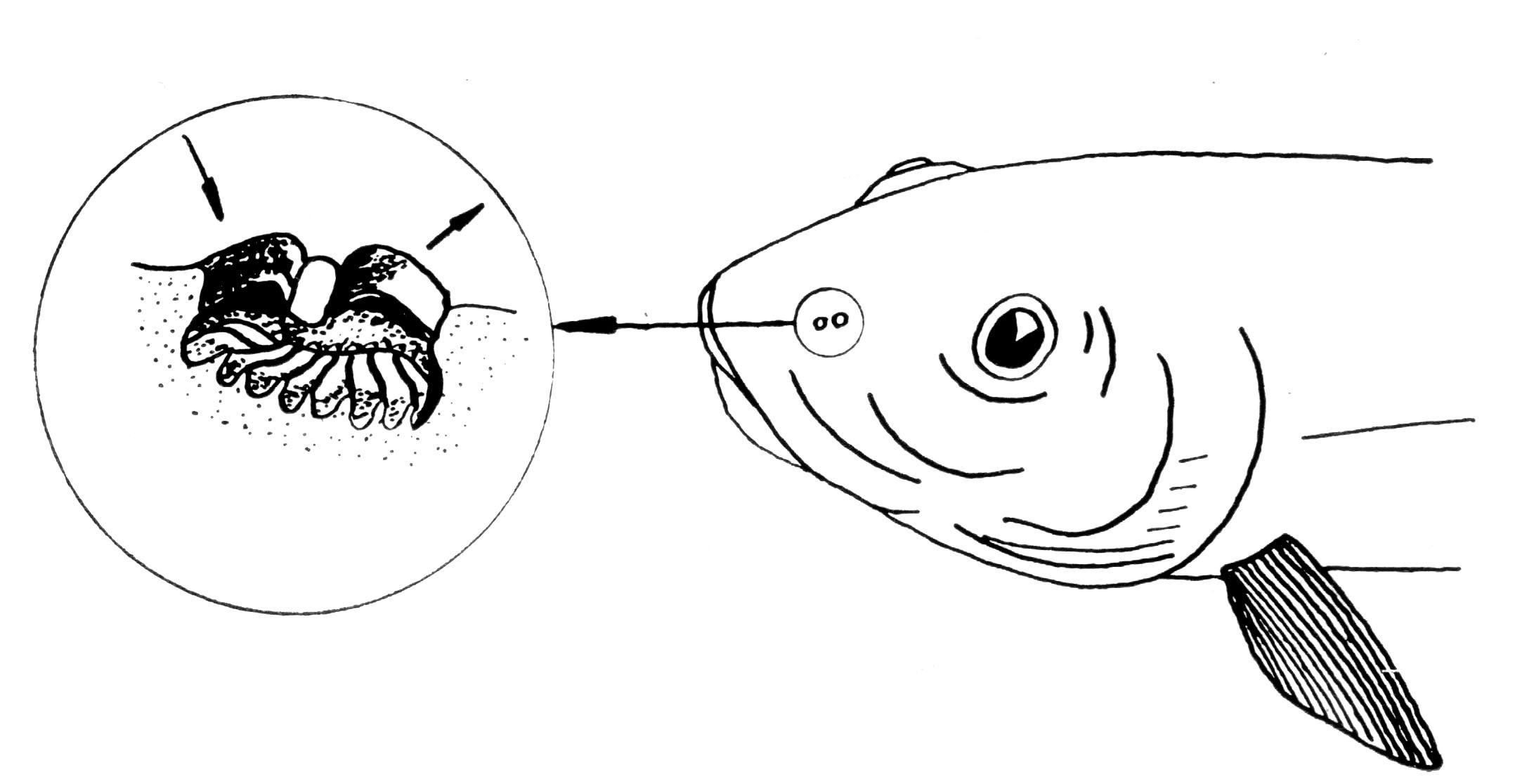 оча
дія хімічних реагентів на нюховий
рецептор відрізняється не спрямованістю,
загальний напрямок на джерело запаху
риба може визначити або за градієнтом
концентрації, або, при рівній концентрації
запаху в навколишньому середовищі, за
спрямованістю потоку води із запаховою
речовиною щодо положення риби. Швидкість
роботи органу нюху набагато інертніша,
ніж зору і слуху (мал. 4.4).
оча
дія хімічних реагентів на нюховий
рецептор відрізняється не спрямованістю,
загальний напрямок на джерело запаху
риба може визначити або за градієнтом
концентрації, або, при рівній концентрації
запаху в навколишньому середовищі, за
спрямованістю потоку води із запаховою
речовиною щодо положення риби. Швидкість
роботи органу нюху набагато інертніша,
ніж зору і слуху (мал. 4.4).
Мал. 4.4. Орган нюху риб
![]() Екстракт
риб’ячого білкового концентрату
очевидно стимулює харчову поведінку,
привабливу спроможність мають також
глютамат натрію, екстракт огірка,
концентрат низькомолекулярних азотистих
сполук і ліприн. Рацемати менш ефективні,
ніж L – ізомери. Гранична чутливість до
L –амінокислот або їхніх LD – рацематам
лежить у межах 1 – 0,01 пікомоль. Чутливість
до D-ізомерів нижча на 3 – 4 порядків.
Екстракт
риб’ячого білкового концентрату
очевидно стимулює харчову поведінку,
привабливу спроможність мають також
глютамат натрію, екстракт огірка,
концентрат низькомолекулярних азотистих
сполук і ліприн. Рацемати менш ефективні,
ніж L – ізомери. Гранична чутливість до
L –амінокислот або їхніх LD – рацематам
лежить у межах 1 – 0,01 пікомоль. Чутливість
до D-ізомерів нижча на 3 – 4 порядків.
Нюх грає помітну роль у харчовій поведінці риб. Так, у тріски прояв характерної пошукової поведінки стимулюється природними харчовими запахами і розчинами деяких вільних амінокислот. Дія внутрішньовидових хімічних стимулів викликає уникнення, його інтенсивність залежить від концентрації запаху. Реакція посилюється за стресування риб – донорів запаху, що має сигнальне значення при виявленні небезпеки. Аналогічно впливають на тріску запахи інших видів біломорських риб – наваги, керчака, камбали. Таким чином, хімічна сигналізація бере участь в регуляції щільності риб на кормових ділянках, забезпечуючи більш раціональне використання кормових ресурсів. За наявності тільки зорового зв'язку між особинами має місце притягнення, тоді як спільна дія зорового і нюхового стимулів викликає реакцію втечі. Це свідчить про головну роль хеморецепції у внутрішньовидових взаємовідносинах під час відгодівлі риб. У осінній період у тріски відбувається зміна екологічної ніші, змінюється її фізіологічний стан. При цьому інтенсивність відповідей на видові запахи помітно знижується, а роль зорових сигналів зростає, що посилює стайні контакти між особинами.
Молодь спроможна сприймати метаболіти хижаків, уникаючи місць їхньої концентрації і формуючи захисні зграї різноманітної конфігурації. Захисна реакція є домінуючою серед усіх інших форм поведінки, наприклад харчової і статевої.
У період нересту територіальні самці коттоїдних використовують хімічні атрактанти для притягнення статевозрілих самиць; феромони самиць стимулюють статеву поведінку самців. Статевозрілий омуль залучається до місць нересту, орієнтуючись на атрактанти, що виділяються зі статевими продуктами. Феромони самок залучають самців і викликають у них усі елементи статевої поведінки.
Високорозвинені системи хеморецепції слугують для риб найважливішими каналами інформації про навколишнє середовище. Хімічні сигнали реалізерної дії викликають у риб прояв різноманітних поведінкових актів, сигнали праймерної дії змінюють фізіологічний статус особини.
Сприйняття запахових подразників у риб, так саме як у наземних хребетних, не чисто ольфакторна, а ольфакто-тригемінальна рецепція. Взаємодія двох систем забезпечує полісенсорне сприйняття зовнішнього впливу. Для риб така полісенсорність нюхової рецепції повинна мати особливу екологічну значимість у зв'язку із водним середовищем мешкання.