

Розділ 16. Стрес і адаптація Стpесоpні реакції
Стрес – сукупність усіх неспецифічних змін, які виникають в організмі під впливом будь-яких факторів, що включають, зокрема, стереотипний комплекс неспецифічних захисно-пристосувальних реакцій (Г. Сельє, 1936 – 1952). Агент, що викликає стрес, називається стресором. Стресором є будь-який достатньо сильний (не обов'язково екстремальний) фактор – тепло, холод, болючі подразники, голодування, інтоксикація тощо. Комплекс неспецифічних захисно-пристосувальних реакцій на базі стресу, спрямованих на створення усталеності (резистентності) організму до будь-якого фактора, позначається Сельє як загальний (генералізований) адаптаційний синдром.
У динаміці загального адаптаційного синдрому закономірно прослідковується три стадії, що характеризують резистентність організму за розвитку стресу: 1) стадія тривоги, 2) стадія резистентності, 3) стадія виснаження. Адаптаційному синдрому передує первинний шок. Тривалість і виразність кожної стадії може варіювати в залежності від природи і сили стресорного агента, виду тварини і фізіологічного стану організму.
Перша стадія синдрому – реакція тривоги – яка протікає гостро, характеризує активну мобілізацію адаптивних процесів в організмі у відповідь на будь-який зсув гомеостазу за стресу. У цей період усталеність організму до несприятливих впливів швидко зростає.
В другій стадії – стадії резистентності – установлюється підвищена опірність до стресора, що носить загальний, неспецифічний характер. Так, наприклад, якщо стрес викликається холодом, то на стадії резистентності виявляється підвищена усталеність не тільки до холоду, але і до інших факторів – підвищеній температурі, токсинів тощо.
У випадках, коли дія стресора тривала, захисні пристосувальні механізми організму можуть виснажуватися і загальний адаптаційний синдром переходити у третю стадію – стадію виснаження, яка характеризується зниженням резистентності організму до даного стресора й інших видів стресорних впливів. Ця стадія називається також повторним шоком. Організатором реалізації загального адаптаційного синдрому в організмі є гіпоталамо – гіпофізарно – надниркова система. Поняття стресу було введено Г.Сельє в 1950 році на підставі досліджень, проведених на ссавцях. Стресорні реакції в риб почали вивчати в 70-х роках, але розуміння перебігу реакцій, які відбуваються у різноманітних умовах і в різних видів риб, поки ще не настільки повне як у ссавців.
При стресі в риб навіть на досить ранніх стадіях можна спостерігати зниження резистентності до хвороб. На стадії виснаження риби знаходяться у важкому фізіологічному стані, наприклад, тихоокеанські лососі після нересту, гинуть, знаходячись у стані, що можна назвати загальним фізіологічним виснаженням. Взагалі в тих випадках, коли хронічний стрес перевищує межі толерантності, починається ослаблення організму, про що часто свідчить спалах захворювання. На початку дії на риб стрес-фактору спостерігається посилення активного і пасивного транспорту іонів натрію і калію через мембрани клітин. За адаптації обидва показники повертаються до достресового рівня, а перед загибеллю активна компонента слабшає, а пасивна – посилюється.
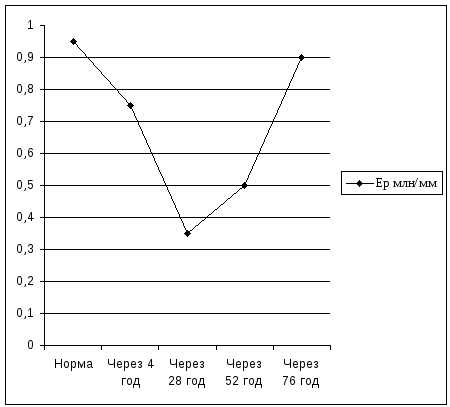
Мал. 16.1. Зміна вмісту еритроцитів за розвитку стрес-реакції
Стресорні реакції викликаються в основному катехоламінами і кортизолом, що діють протягом двох різних, але перекриваючих один одного відрізків часу. Зміни, викликані в організмі катехоламінами (адреналіном і норадреналіном), відбуваються менше ніж через секунду і тривають від декількох хвилин до годин, ймовірно, під контролем із боку симпатичної нервової системи. Кортизол викликає зміни, які починаються менше ніж через годину і тривають тижні і навіть місяці. Симпатична нервова система виділяє катехоламіни швидко і недовго, у той час як хромафінові клітини в головній нирці можуть продукувати їх протягом тривалого часу. У риб кортизол утворюється інтерреналовою тканиною в головній нирці під впливом АКТГ гіпофіза. Утворення кортизолу починається не раніше ніж через 15 хв., але для досягнення максимального рівня продукції гормону потрібно біля 24 годин.
Спільний вплив катехоламінів може бути схарактеризовано як підготовка до термінових дій. Вони викликають підвищену рухливість (занепокоєння, реактивність), мобілізацію енергетичних запасів, посилення дихання й інші функціональні зміни.
Таблиця
16.1. Зміни частоти дихання у риб після
витягування із води на 5 і 10 хв. при t 4°C![]()
|
Дія подразника, хв. |
Частота дихання до подразнення |
Частота дихання після подразнення |
Частота кашлю |
|
5 |
33,0±0,7 |
39,0±0,8 |
6,6±0,6 |
|
10 |
33,4±0,5 |
44,4±1,1 |
11,2±0,8 |
Специфічні результати, що викликаються підвищеним вмістом катехоламінів, наведені в таблиці 16.2.
Таблиця 16.2. Вплив підвищеного вмісту катехоламінів у риб
|
Результат |
Примітки |
|
1. Збільшення вмісту глюкози в крові |
1. При нормальному вмісті 50 - 150 мг/ 100 мл, вміст глюкози збільшується до 200 - 300 мг/100 мл. |
|
2. Збільшення частоти серцебиття |
2. Ін'єкція адреналіну може майже подвоїти частоту серцебиття. |
|
3. Збільшення хвилинного об'єму серця |
3. Може відбуватися в основному в результаті збільшення ударного об’єму і незначного зростання частоти серцебиття. |
|
4. Збільшення дихальної вентиляції |
4. Звичайно супроводжується зменшенням кількості доступного кисню, що дифундує із води. |
|
5. Розширення і звуження артеріол |
5. У ссавців (а можливо і в риб) звуження одних артеріол і розширення інших призводить насамперед до збільшення хвилинного об'єму серця для переносу крові до голови, серця і скелетних м'язів, але не до внутрішніх органів. |
|
6. Підвищення вмісту тромбоцитів в крові |
Мабуть, через скорочення селезінки. |
|
7. Погіршення перистальтики або загальний стаз кишечнику |
У риб у стані стресу не спостерігалося, але відзначено в ссавців. Збільшується число спостережень, що припускають наявність цього явища в риб. |
|
8. Розщеплення глікогену до глюкози в печінці і м'язах; звільнення резервних ліпідів |
8. Ці речовини слугують джерелом енергії для забезпечення підвищеного рівня активності. |
Необхідна кількість катехоламінів для прояву цих впливів майже невідома, оскільки хімічне визначення кількості адреналіну і норадреналіну важке і потребує великих об'ємів плазми крові через їхні низькі концентрації. У дорослих особин, що повернулися на нерестовища, у спокійному стані вміст катехоламінів у плазмі складає біля 1 мкг/ мл, а після 25 – хвилинного стресу (рибу витримували без води) – біля 20 мкг/ мл. Загальна концентрація адреналіну в райдужної форелі в спокійному стані збільшується приблизно із 0,007 до 0,160 мкг/мл через годину після стресу, що продовжувався 10 хв. Ці два спостереження об'єднує тільки те, що вміст адреналіну в крові зростає під час стресу. Отримані результати настільки відмінні один від одного, що їх важко зіставити із даними для ссавців або робити на їхній основі які-небудь узагальнення. Співвідношення адреналіну і норадреналіну в риб значно варіює. У ссавців катехоламіни на 70 % представлені норадреналіном, те ж саме у коропа. У кижуча адреналіноподібні речовини приблизно на 70 % подані адреналіном, а в деяких інших риб адреналіну і норадреналіну утримується порівну.
Кортизол впливає на багато систем органів. На відміну від катехоламінів, що, очевидно, сприяють виживанню, кортизол, особливо за тривалого стресу, часто несприятливо діє на організм. Спочатку вплив виявляється на ділянках, де відбувається іонний обмін: зяброві мембрани, ниркові канальці, сечовий міхур і епітелій кишечнику (в риб у морській воді). На цих ділянках кортизол, певно, є антагоністом пролактину, оскільки він збільшує проникність мембран для іонів.
|
Органи - мішені або функції |
Дія |
|
Загальні функції
білковий обмін
вуглеводний обмін
кровообіг
У прісній воді зябра |
Прискорений синтез білків через підвищену активність РНК; синтезуються, певне, в основному протеолітичні білки Пригнічення росту Мобілізація білка із тимуса, селезінки і печінки
Уповільнена утилізація вуглеводів Прискорене утворення глюкози із білків тканин Відкладення глікогену в печінці
Мобілізація лейкоцитів
Ослаблена запальна реакція |
В умовах ставкового, а тим більш, садкового розведення риб рибники повинні дуже пильно відноситися до проведення технологічних заходів вживаючі заходи, щоб знизити стресуванні риби. Такі технологічні заходи як облов, транспортування тощо призводять до значного стресування риби, що проявляється в збільшенні виділення адреналіну та стероїдних гормонів. Все це негативно впливає на всі фізіологічні параметри рибі та знижує її резистентність. Оцінку стану риби можливо давати, виходячи із картини крові та гормонального профілю риби. Далі ми даємо декілька прикладів тих змін, які відбуваються у риб при несприятливих умовах.
Нерестові міграції
Нерестова міграція тихоокеанських лососів як реакція на стрес, уперше досліджувана Робертсоном із співробітниками на чавичі із річці Сакраменто. У чавичі, пійманої в морі на узбережжі Сан-Франциско, вміст кортизолу був низьким, але збільшувався в 4 – 5 разів через 24 години після поміщення її в ємності на борту дослідницького судна. Було також виявлено підвищення основних рівнів кортизолу в риб із зростанням шляху, пройденого до верхів'я по річці. На нерестовищах досягалося максимального рівня кортизолу, подібне тому, яке було в риб, спійманих у морі та поміщених у ємкості. В міру проходження міграційного шляху в риб досягаються піки рівнів кортизолу, близькі до тих, які спостерігаються після вилову риб.
Підвищений катаболізм білків, що стимулюється кортизолом, у кінцевому рахунку торкається і білки плазми крові, що включають і білкові антитіла. Знижений вміст антитіл обумовлює підвищену схильність до хвороб. У тихоокеанських лососів, які йдуть на нерест, розвивається звичайно хвороботворний цвілевий грибок Saprolegnia, що розростається у вигляді шкірки або губки на тілі риб і руйнує шкіру і м'ясо. Проте на противагу втраті антитіл із білків плазми крові частота серцевих скорочень і тиск крові в горбуші, що завершили нерест і гинуть, зберігається майже нормальними – серце і кровоносні судини відносяться до органів, що відказують в останню чергу.
Зовсім не обов'язково, щоб тихоокеанські лососі гинули після завершення нересту, і більшість інших лососевих, безсумнівно, розмножуються декілька разів. Коли нерки нерестилися в штучно контрольованих умовах і утримувалися після цього в знезараженій прісній воді, вони виживали в ній до 11 міс. Приблизно через 2 місяців після нересту вони починали харчуватися і до них повернулося властиве морському періоду життя зелене і сріблясте забарвлення. Загнуті щелепи, проте, залишилися, а спроможність до осморегуляції в морській воді була досліджена. Можна припустити можливість переходу дорослих особин, що віднерестилися, до морського способу життя. Виявлено, що в сталевоголового лосося в процесі нерестової міграції відбувається накопичення жиру (атеросклероз, рання стадія затвердіння артерій) у коронарних артеріях. Ці явища зникають після повернення в морську воду. Хоча біля третини сталевоголових лососів, що нерестяться, гинуть (включаючи промислову смертність), міграційний стрес у цього виду, очевидно в основному менш сильний, ніж у тихоокеанських лососів, – тіло менш виснажене, щелепи скривлені не настільки помітно і т. ін. У атлантичного лосося реакція на міграційний стрес приблизно така ж як у сталевоголового.
Таким чином, нерестова міграція в лососевих є сильним стресом (звичайно летальним) і роль кортизолу в ньому установлена достатньо точно. Роль глюкози і катехоламінів поки не вивчена, але, очевидно, вона достатньо складна через надмірну м'язову напругу і зміни в тканинах за тривалого голодування.