

Спеціальна фізіологія центральної нервової системи Структура і функції спинного мозку
Спинний мозок риб знаходиться в спинномозковому каналі, уздовж якого розташовуються нервові клітини переважно мультиполярного типу (з великою кількістю відростків). У передній частині тіла він переходить у довгастий мозок. Спинний мозок вкритий трьома мозковими оболонками: м'якою або судинною (внутрішньою), павутинною (середньою) і твердою (зовнішньою) Простір між м'якою і павутинною оболонками (підпавутинний простір) і спинномозковий канал заповнені спинномозковою рідиною.
У круглоротих спинний мозок має вигляд плоскої стрічки, що складається із боку черевної порожнини із сірої речовини, в якій містяться тіла нервових клітин, а у верхній білій частині знаходяться провідні елементи. Починаючи із круглоротих, намічається найважливіший процес утворення довгих висхідних провідних шляхів спинного мозку, що є основою цефалізації. У кісткових риб на розрізі спинний мозок має округло-трикутну форму. Сіра речовина утворює дорзальні і вентральні роги; на периферії знаходиться біла речовина, що включає дорзальні, вентральні і латеральні стовпи (мал. 3.4).
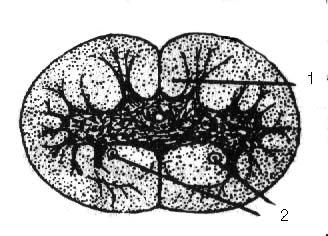
Мал. 3.4. Поперечний зріз спинного мозку
1 – провідні шляхи; 2 – маутнеровські волокна
Дорзальні стовпи складаються із аферентних нервових волокон, що направляються до головного мозку. У риб провідні шляхи бічних стовпів доходять до довгастого мозку і мозочка, надалі у філогенезі вони доходять до середнього мозку і, нарешті, у ссавців доходять до таламічної ділянки. У хрящових і кісткових риб формуються такі важливі для хребетних тварин висхідні шляхи, які зв’язують спинний мозок із найважливішими апаратами центральної регуляції руху: вестибулярним апаратом і мозочком, а також відповідні низхідні (вестибулярно – спинальний шлях, церебело (мозочково) – спинальний шлях).
Рухові волокна, що ідуть від головного мозку знаходяться у вентральних стовпах, а латеральні стовпи складаються із відцентрових і доцентрових волокон.
Спинний мозок функціонально поділяється на сегменти відповідно кількості хребців, від яких відходять спинномозкові нерви. Вони виходять двома корінцями – вентральними (черевними) і дорзальними (спинними). Дорзальні корінці беруть початок від біполярних нервових клітин, розташованих у спинномозкових гангліях, цими корінцями збудження йде у спинний мозок. Вентральні корінці починаються від моторних клітин, розташованих у вентральних рогах, цими корінцями збудження поширюється на периферію. До складу вентральних корінців входять волокна вегетативної нервової системи. У ланцетника і міног спинні і черевні корінці не з'єднуються й утворюють самостійні нерви. У міксин і риб вентральні і дорзальні корінці з'єднуються після виходу зі спинного мозку й утворюють змішані (несучі аферентні, еферентні і вегетативні волокна) спинномозкові нерви, що виходять посегментно із правого і лівого боків тіла. Спинномозковий нерв далі розпадається на три гілки – спинну, черевну і внутрішню, що іннервують відповідні м'язи і кровоносні судини, а також внутрішні органи. До складу внутрішньої (вісцеральної) гілки входять волокна вегетативної нервової системи.
Під час перерізування дорзальних корінців спостерігається порушення чутливості в сегментах, що відповідають перерізаним корінцям, під час перерізування вентральних корінців спостерігаються рухові збудження у відповідних міомерах. Якщо ж переріз зробити тільки в одному сегменті, то ніяких порушень не відбудеться, тому що кожний спинномозковий нерв іннервує не тільки відповідний йому, але і прилеглі до нього попереду і позаду метамери. Це вказує на перекриття зон поширення аферентних і еферентних нервових волокон. Під час подразнень метамерів тіла виникають біоструми в сусідніх дорзальних корінцях.
Спинний мозок виконує рефлекторну і провідникову функції.
Рефлекторна функціяполягає в здійсненні спинним мозком деяких простих рефлексів. Нерви, що йдуть від нього, іннервують усі скелетні м'язи, за винятком м'язів голови, іннервацію яких здійснюють черепно-мозкові нерви. Спинний мозок бере участь у виконанні різноманітних рухових реакцій тулуба і плавців. У деяких риб сильний розвиток периферичних нервів призводить до збільшення зон спинного мозку відповідно центрам цих нервів. Так, у морського півня розвинені п'ять парних потовщень вентрального відділу передньої частини спинного мозку відповідно п'ятьом нервам, що іннервують передні збільшені і спроможні до самостійних рухів промені грудних плавців.
Деякі реакції здійснюються тільки завдяки центрам спинного мозку, інші – завдяки різноманітних відділам головного мозку, в останньому випадку спинний мозок виконує провідникову функцію.
У спинному мозку беруть початок еферентні нейрони, що регулюють роботу внутрішніх органів, – судинорухові центри, центри які керують хроматофорами та ін. У риб, що мають електричні органи, в певних його сегментах є особливі гангліозні клітини, що дають початок спеціальним нервовим волокнам, які направляються до електричних органів. У тому випадку, коли електричні органи мають м'язове походження, нерви електричних органів починаються в моторних клітинах.
Провідникова функціяспинного мозку полягає в проведенні збудження як у висхідному напрямку – до головного мозку, так і в низхідному – від головного мозку через спинний по спинномозкових нервах до м'язів і різних органів. У спинний мозок надходять імпульси від різноманітних рецепторів – пропріорецепторів м'язів, суглобів, зв’язок, інтеро- і екстерорецепторів. Ця інформація (аферентні імпульси) по бічних висхідних рогах передається у довгастий мозок і мозочок.
Проведення імпульсів волокнами провідних трактів здійснюються із різною швидкістю. Так у коропа швидше (58 – 63 м/с) проводяться тактильні сигнали, вентрально-латеральні стовпи проводять спадні моторні імпульси зі швидкістю 18 – 20 м/с; у ретикуло-спинальних волокнах швидкість проведення найменша.
Серед еферентних елементів у спинному мозку риб слід зазначити гігантські аксони маутнеровських клітин – двох великих нейронів, розташованих в ділянці довгастого мозку, на рівні ядер 8-ої пари нервів. Їх нервові волокна утворюють ретикуло-спинальний шлях, який поширюється до кінця спинного мозку. Ці клітини відіграють роль у запуску стартового ривка риби, викликаючи різкий рух хвостового плавця. Вони відсутні або погано розвинені в донних риб або риб, що мають інший характер локомоції.
Якщо порушити зв'язок відділів головного мозку, що лежать вище, із довгастим, але зберегти зв'язок довгастого і спинного мозку, у риб зберігається спроможність до пересування, але порушується координація рухів. У разі відділення спинного мозку від довгастого наступає явище шоку – тварина не здатна рухатися. Шок можна пояснити тим, що порушення зв'язку припиняє потік імпульсів від головного мозку, що регулює роботу спинного мозку і його центрів, розташованих нижче місця перерізу, котрі якийсь час не спроможні до самостійної роботи. Велике значення за цього має вимикання імпульсів від ретикулярної формації мозкового стовбура, які в нормі активують роботу спинного мозку. Чим вище рівень, на якому знаходиться тварина в еволюційному розвитку, тим довше у неї продовжується шок. Так, у риб і амфібій шок триває 2 – 15 хв., у птахів – декілька годин, у собак – декілька тижнів, у людини – декілька місяців. Коли проходить шок, відновляються тільки рефлекторні рухи, тобто відповідні реакції на будь-які зовнішні подразнення. Наприклад, здригування у відповідь на уколювання голкою.
У риб, на відміну від вищих тварин, спинний мозок регенерує і його діяльність відновляється. Гістологічні дослідження виявляють регенерацію нервових шляхів. У акул із перерізаним спинним мозком спостерігаються не тільки рухи передньої частини тіла, але й узгоджені із нею рухи задньої частини, що розташовані нижче місця перерізу. Це пояснюється тим, що передня частина тіла рухається за рахунок того, що рухові центри передньої частини спинного мозку збережені. Рухи цих м'язів механічно передаються м'язам задньої частини тіла, тому що хвиля м'язового скорочення проходить по всьому тулубі. У пропріорецепторах м'язів задньої половини виникає збудження, що передається у відповідні сегменти спинного мозку до рухових центрів, а звідти – знову до м'язів. Таким чином, для існування локомоторного ритму в спинальних риб потрібна аферентна сигналізація. Якщо перерізати дорзальні, аферентні корінці або зруйнувати задні сегменти спинного мозку, то рухові реакції зникнуть.
Характерною рисою спинного мозку риб є наявність у ньому нейросекреторних елементів, що мають багате кровопостачання, так, на кожну нейросекреторну клітину спинного мозку ската припадає від 4 до 5 капілярів. Ці клітини переважно знаходяться в каудальної частині спинного мозку, образуючі урофіз, який приймає участь в осморегуляції .