

Мал. 9.3 Електрокардіограма севрюги
Електрокардіограма риб має всі характерні зубці, які виявляються під час запису струмів дії серця людини. Під час скорочення передсердя записується зубець Р, а під час скорочення шлуночка зубці QRST. Зубцю Р у риб передує зубець V, що формується в області венозного синуса, а у пластинчатозябрових на ЕКГ є зубець U. Він відбиває процес деполяризації у артеріальному конусі і записується між зубцями S і T (мал.9.3).
Число серцевих скорочень. Число серцевих скорочень у риб приблизно дорівнює числу дихальних рухів і залежить від багатьох факторів. Зокрема, число скорочень залежить від температури: чим вище температура, тим більше число серцевих скорочень. Залежність серцевих скорочень (ЧСС) ізольованого серця строкатого товстолобика від температури середовища наведена у таблиці 9.1.
Таблиця 9.1. Частота скорочень ізольованого серця
|
Температура, С |
ЧСС |
|
0 |
6 |
|
5 |
9 |
|
10 |
10 |
|
15 |
13 |
|
20 |
19 |
|
25 |
21 |
|
30 |
24 |
Під час руху ритм серця пришвидшується, поверненню крові сприяє насосна функція хвостових м’язів. На частоту серцевих скорочень впливає і вміст аміаку у воді (під його дією частота серцевих скорочень знижується тим більше, чим більше аміаку у воді) та інші фактори.
Хвилинний об’єм серця – це об’єм крові, що виштовхується шлуночком на протязі 1 хв. Цей показник залежить від багатьох факторів, у тому числі і від швидкості плавання. Так, у форелі масою 1000 г при 10 С в спокою хвилинний об’єм складав 12 мл, при плаванні зі швидкістю 75% від критичної – 21 мл і 95% – 32 мл. У звичайних колючих акул масою 2,3; 3,5 і 6,1 кг хвилинний об’єм серця дорівнював 41, 23 і 16 мл/хв*кг).
У таблиці 9.2 приведені величини хвилинного об’єму серця деяких риб.
Таблиця 9.2. Хвилинний об’єм серця риб
|
Назва риби |
Частота скорочень серця, уд /хв. |
Відносний хвилинний об’єм, мл/кг |
|
Тріска |
30 |
9,3 |
|
Сазан |
16 |
18,7 |
|
Райдужна форель, 1200 г, 10С |
38 |
18 |
|
Нерка 1000 г, 5С, спокій |
31 |
21 |
|
Нерка 1000 г, 5С, плавання |
47 |
65 |
|
Нерка 1000 г, 15С, спокій |
52 |
31 |
|
Нерка 1000 г, 15С, плавання |
83 |
93 |
|
Нерка 1000 г, 22С, спокій |
86 |
64 |
|
Нерка 1000 г, 22С, плавання |
105 |
103 |
У дослідженнях, проведених на знерухомлених рибах, було показано, що хвилинний об'єм серця у пластинчатозябрових риб більше, ніж у костистих. У коропа (без наркозу) хвилинний об'єм складав 18,3 мл/кг, а у акули він досягав 22 мл/кг.
Робота, що виконується серцем (W), визначається рівнянням
![]()
де Р - систолічний тиск, а SV - ударний об’єм серця.
Фізіологія кровоносних судин
Кровоносна система хребетних тварин замкнена. Кров рухається кровоносними судинами, які являють собою трубки різного діаметра.
Функціональні групи судин. Судини кровоносної системи мають різну будову і різне функціональне призначення. У залежності від виконуваної ними функції судини підрозділяються на 6 груп: 1) амортизуючі; 2) судини опору; 3) судини - сфінктери; 4) обмінні; 5) ємнісні; 6) шунтуючі.
Амортизуючі судини – це найбільш значні магістральні артерії, у котрих ритмічно пульсуючий, мінливий потік крові перетворюється у більш рівномірний і плавний. Стінки цих судин містять мало м’язових елементів і багато еластичних волокон.
Судини опору –(резистивні судини) діляться на передкапілярні (дрібні артерії й артеріоли) і посткапілярні (венули і дрібні вени). Співвідношення між тонусом пре- та посткапілярних судин визначає рівень гідростатичного тиску у капілярах, величину фільтраційного тиску й інтенсивність обміну рідини. Резистивні судини вміщують невеличку частину крові, але на 2/3 створюють загальний периферичний опір.
Судини – сфінктери – це останні відділи передкапілярних артеріол. Їхній тонус забезпечує зміну числа функціонуючих капілярів, тобто забезпечує зміну кількості крові, що протікає через певну ділянку.
Обмінні судини – капіляри, у яких відбувається обмін речовинами між кров'ю і тканинами. Стінки капілярів не містять м’язових елементів і не спроможні до скорочення.
Ємнісні судини –складають венозний відділ серцево-судинної системи. Створюючи лише малу частку периферичного опору, вони вміщують 3/4 усієї крові. Деякі вени мають особливу ємність як депо крові.
Шунтуючі судини – це артеріовенозні анастомози, що забезпечують прямий зв'язок між дрібними артеріями і венами в обхід капілярної мережі. Коли ці судини відкриті, кровотік через капіляри або зменшується або припиняється.
Рух крові в судинах підпорядковується законам гідравліки. У такій системі рідина рухається від місця, де спостерігається більший тиск до меншого; плин рідини забезпечується різницею тисків. Якщо ми побудуємо модель такої системи – з'єднаємо бак із довгою трубкою і по всій довжині її встановимо манометри для визначення тиску рідини, то зауважимо, що в міру просування до вихідного отвору тиск у системі падає. Тиск при цьому витрачається на:
1. тертя: а) зовнішнє – тертя рідини об стінки трубки і;
б) внутрішнє – тертя часток одна із одною;
2. менша частина тиску витрачається на надання швидкості поточній рідині.
Якщо просвіток трубки буде неоднаковий, а місцями буде звужуватися, як у кровоносній системі, то при проходженні через звужену ділянку тиск буде різко падати. Він буде витрачений на подолання опору на вузькій ділянці.
Якщо у систему включені еластичні трубки, то частина тиску витрачається на розтяг стінок цих трубок, переходячи у пружну потенційну енергію. Отже, у кровоносній системі тиск крові обумовлений роботою серця і буде падати у міру віддалення від серця. Крім того, чим більше будуть розгалужуватися судини, тим більше буде опір токові крові і тим значніше буде падати тиск.
Тиск у різних ділянках кровоносної системи залежить від типу судин і відстані від серця. У райдужної форелі у спокої тиск у черевній аорті складає 40/32 мм, а в спинній аорті 29/25 мм ртутного стовпа. Під час руху венозний тиск досягає 9 мм рт. ст., тиск у черевній аорті зростає на 40 %, а у спинній – на 16 %. У акули черевна аорта містить вдвічі більше еластину, ніж спинна. Еластин має набагато більш низький модуль пругкості, ніж колаген, тому черевна аорта порівняно ригідна, і при підвищенні тиску потік крові через зябра зростає мінімально.
Таблиця 9.3. Величина тиску крові в судинах
|
Назва риби |
Судина |
Артеріальний тиск, мм рт. ст. систолічний /діастолічний |
|
Лосось |
Черевна аорта |
40/32 |
|
Лосось |
Спинна аорта |
29/25 |
|
Тихоокеанський лосось |
Спинна аорта |
44/38 |
|
Тріска |
Луковиця аорти |
29/18 |
|
Колюча акула |
Черевна аорта |
30/24 |
|
Колюча акула |
Спинна аорта |
17/16 |
|
Скат |
Черевна аорта |
16/14 |
|
Сом |
Черевна аорта |
40/30 |
Тиск, який утворюється серцем кісткових риб, вище, ніж у хрящових близької маси, що добре узгоджується із різницею у величині серця. Величина напірного тиску у аорті знаходиться у прямій залежності від розмірів тіла. Так, середній тиск у спинній аорті у звичайної колючої акули довжиною 30, 60 і 90 см дорівнював відповідно 15, 26 і 35 мм рт. ст. Від 20 до 70% напірного тиску витрачається на кровотік через зяброве русло (мал.9.4).
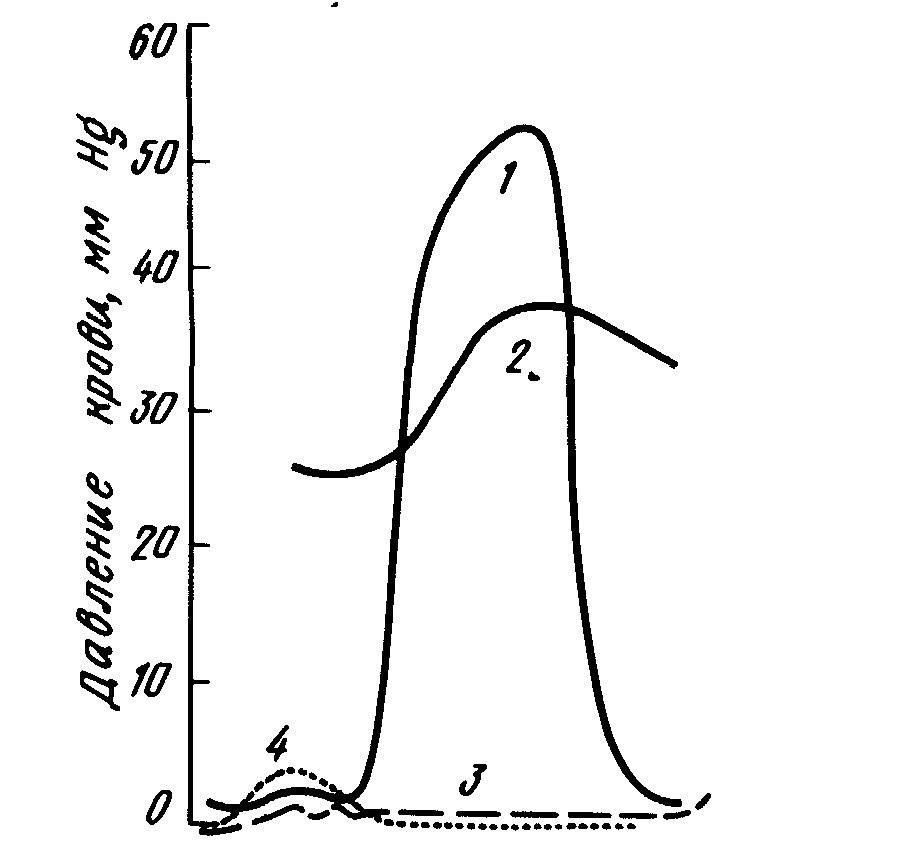
Мал. 9.4. Тиск крові у різних відділах серця райдужної форелі під час циклу скорочення: 1 - шлуночок; 2 - цибулина аорти; 3 - венозний синус; 4 - передсердя
Кровоносне русло у зябрах схоже за своєю будовою на кровоносне русло у легенях наземних хребетних. Як і у легенях, у зябрах є щільна мережа капілярів, що займає 60 – 80 % поверхні зябрової пластинки (біля 4 м довжини капілярів на 1 см2). Щільність зябрових капілярів, як і їхня загальна протяжність, залежать від віку і величини риби, а також від умов існування. Так, у придонних в'юнів поверхня зябрових капілярів (см2 на 1 г маси тварини) складає 5,3, а у судаків 18,3.
У риб, що постійно живуть в умовах дефіциту кисню, розвивається шкірне дихання. Воно може досягати 20 – 30% загального дихання і вище у молодих риб. Як правило, це риби, що ведуть придонний спосіб життя і зариваються у ґрунт або мул. У них тонкий, часто позбавлений луски, багато васкуляризований шкірний покрив. Так, у звичайних в'юнів на 1 см2 шкіри є 3,1 м капілярів, їхня поверхня складає 1,5 см2 на 1 г маси (поверхня зябрових капілярів – 5,3 см2/м). У в'юнів в диханні бере участь і багато васкуляризована ділянка кишки.
Найбільша щільність шкірних капілярів – 4,5 м/см2 виявлена у білокровних риб, що не мають у крові еритроцитів. При дуже низькій кисневій ємності крові в цих риб велика поверхня зябер і низький опір току води через них. Наприклад, у білокровки масою тіла 770 г киснева ємність 0,7 мл О2/100 мл крові, поверхня зябер - 4.2 см2/г, а їхній опір у 5 разів нижче, ніж у донної риби морського чорта (1,55 кг) із кисневою ємністю крові 6,5 мл О2, і поверхнею зябер 1.4 см2/г. У білокровок велику участь у диханні приймають капіляри, які розташовані у шкірі і плавцях, сумарна поверхня яких у 3 рази більше поверхні тих, які розташовані у зябрових пластинках. Відмічено, що щільність капілярів у шкірі риб із віком змінюється мало. Тому у молодих маленьких риб із відносно великою поверхнею шкіри протяжність розташованих у ній капілярів у розрахунку на 1 г маси тіла вище, ніж у старих великих риб.
Про те як розподіляється у риб артеріальна кров після проходження через зябра дозволяють судити дослідження, проведені на сибірському харіусі. Доведено, що у харіуса більш 60% хвилинного об'єму серця протікає через скелетні м'язи, причому майже чверть цієї кількості адресована червоним м'язам, об'єм яких дуже малий. Ці м'язи у риб виконують вкрай важливу функцію, забезпечуючи необхідний комплекс рухів для плавання, поворотів і дихання: білі м'язи беруть участь тільки в епізодичних ривкових рухах. Тому не випадково «спочиваюча» рухова активність червоних м'язів значно вища, ніж білих. Природно, що у червоних м'язах харіуса – швидкохідної риби, що далеко мігрує – виявлений високий кровотік, близький до кровотоку у нирках, печінці і сумірний із м'язовим кровотоком у ссавців тієї ж маси.
В цілому тиск крові у судинах в спокої такий: аортальна цибулина біля 35 мм рт. ст. під час систоли та 25 під час діастоли, у судинах зябер ще відмічаються пульсові хвилі і тиск знижується 10 – 7 мм рт. ст., у судинах тиск підтримується на рівні 17 – 5 мм рт. ст., у капілярах він падає до 0, а у венах до – 15 мм рт. ст. Під час руху тиск значно підвищується.
Таблиця 9.4. Кровопостачання органів сибірського харіуса
|
Показники |
Зябра |
Селезінка |
Печінка |
Нирки |
Гонади |
М'язи | ||
|
білі |
червоні |
усі | ||||||
|
Маса, % до маси тіла |
1,9 |
0, 061 |
0,73 |
0.78 |
2.6 |
55 |
2 |
57 |
|
Частка хвилинного об'єму серця, % |
100 |
1,5 |
11 |
6,1 |
0,18 |
49 |
14 |
63 |
|
Кровотік, мл/хв на 100 г органа |
174 |
81 |
50 |
26 |
0,.23 |
2,9 |
23 |
3,6 |
|
Кількість циркулюючої крові, мл на 1 кг органа |
87 |
221 |
145 |
- |
- |
12 |
30 |
- |
Виявлено прямий зв'язок між швидкістю об'ємного кровотоку у м'язах риби і щільністю розташованих у ній капілярів. Так, у червоних м'язах у порівнянні із білими щільність капілярів набагато вище, середня довжина одного капіляра помітно коротше, а м'язове волокно тонше. У звичайного окуня щільність капілярів у червоних м'язах в 10,3 рази більша, ніж у білих (відповідно 620 і 60 капілярів на 1 мм2), їх довжина в 1,36 разів менша (750 і 1020 мкм), а діаметр червоного волокна у 2,37 разів менший за діаметр білого (53 і 126 мкм). У стерляді, як риби малорухливої, ці показники розрізняються менше – щільність капілярів у червоних м'язах у 4,8 рази більша, ніж у білих (відповідно 595 і 125 капілярів на 1 мм2), їх довжина у 1,68 разів менша (770 і 1280 мкм), а діаметр червоного волокна всього у 1,3 разів менший за діаметр білого (54 і 70 мкм). Всі ці фактори сприяють переносу О2 із крові у м'язове волокно і тому різко покращують умови кровопостачання червоних м'язів. Відмічено, що у червоних м'язах деяких риб завдяки протиточному розташуванню дрібних артерій і вен підтримується більш висока температура тіла, ніж у білих м'язах, що сприяє збільшенню швидкості окисних процесів.
На кровопостачання м'язів риби значний вплив мають і умови існування. Так, у круглих карасів – риб із високим рівнем шкірного дихання, завдяки якому вони спроможні переживати висихання водоймищ, – підвищена васкуляризація шкіри супроводжується низькими показниками кровопостачання скелетних м'язів, що за своїми величинами наближаються до показників кровопостачання м'язів стерляді.
Цікава васкуляризація газової залози плавального міхура риб. Щільність тіла риби у 1,06 разів вища, ніж води; тому у рівноважному стані біля 6% об'єму риби зайнято газом, частіше усього О2, що секретується у порожнину плавального міхура залозою, розташованої у його стінці. Механізм переносу газу із крові в міхур досить складний. Напруга газу у крові риб, на якій би глибині вона не знаходилася, не може перевищувати напругу цього газу у навколишній воді, що у свою чергу близько парціальному тиску газу у атмосфері. Для О2, наприклад, він складає 0,2 атм. У той же час кисень, що знаходиться у плавальному міхурі, відчуває тиск, рівний тиску води над рибою (із зануренням на глибину 10 м тиск збільшується на 1 атм). Переніс О2 із крові до міхура, вбік більш високої напруги кисню здійснюється завдяки особливому розташуванню кровоносних судин у стінці міхура.
Виявилося, що дві третини залози займають кровоносні судини, що утворюють три капілярних русла, послідовно сполучених між собою більш значними магістральними судинами. У капілярах середнього русла, що лежать на внутрішній поверхні міхура, О2 переноситься із крові у порожнину міхура. Два інших русла, названих А. Крогом "чудесною мережею", складаються із довгих капілярів, розташованих паралельно і впереміжку між собою. По артеріальних капілярах кров із спинної аорти тече до міхура, по венозних – повертається назад і направляється у ворітну вену печінки.
Завдяки тісному зіткненню артеріальних і венозних капілярів кисень, що розчинився у великих кількостях у крові, яка відтікає від міхура (пригадаємо, що кількість розчиненого газу в рідині пропорційна тиску цього газу над рідиною, а у міхурі він може бути дуже великим), встигає за час руху крові венозними капілярами “чудесної мережі” перейти у кровартеріальних капілярів і створити там рівноважну концентрацію. Чим вище тиск газів у міхурі, тим більше часу потрібно для створення такої концентрації. Тому у глибоководних риб довжина капілярів досягає 10 – 25 мм. У результаті такої роботи протитічної системи на зовнішніх кінцях “чудесної мережі” концентрація розчиненого у крові артеріальних і венозних капілярів кисню буде однакова, причому на внутрішній поверхні міхура вона відповідає тиску у ньому О2, а на виході із залози – напрузі О2 у воді, що протікає через зябра.
Принцип протитоку дозволяє представити і можливий механізм переносу О2 із крові у порожнину міхура. Такий переніс буде відбуватися, якщо у середньому руслі капілярів газової залози Ро2 стане вище, ніж у порожнині міхура. Надлишок Ро2 виникає в початковій частині венозних капілярів, які несуть кров із міхура, за рахунок дисоціації НbО2 під впливом молочної кислоти, що утворюється у газовій залозі. Звільнений О2 із еритроцитів потрапляє спочатку до плазми крові венозних капілярів, потім у плазму крові артеріальних капілярів. що лежать поруч і із током крові потрапляє у капіляри середньої мережі. Якщо цей ефект повторюється, то у венозних капілярах знову буде з'являтися надлишок розчиненого у плазмі О2 порівняно із О2 у плазмі артеріальних капілярів і який буде дифундувати у міхур, збільшуючи у ньому концентрацію і тиск кисню.
Отже, у риб велика різниця у артеріальному постачанні кров’ю різних органів і тканин. Завдяки цьому при малому хвилинному об’ємі серця у життєво важливих органах, у тому числі органах руху, постійно підтримується високий кровотік. Далі, риби мають можливість розвивати додаткові дихальні русла у шкірі і, частково, у кишечнику, що дуже важливо при існуванні за умов дефіциту О2. Отже, посилення шкірного дихання супроводжується деяким перерозподілом артеріального кровотоку із зниженням його у скелетних м’язах (риби із розвиненим шкірним диханням, як правило, менш рухливі). Крім того, у більшості риб на відміну від інших тварин у плавальному міхурі є значний резерв О2, користуватися котрим риба може при зміні глибини занурення. Нарешті, у риб – водних тварин – енергетична вартість процесів, які підтримують форму тіла і руху, нижча, ніж у наземних тварин.