

Система залоз внутрішньої секреції
Система залоз внутрішньої секреції риб складається із значної кількості залоз різної співпідпорядкованості, які знаходяться у певних співвідношеннях із нервовою системою. По-перше, це залози, які або підпорядковані нервовій системі, або є нервовими клітинами, для яких властива нейросекреція. По-друге, це залози, які регулюються певними гормонами гіпофізу і, по-третє, залози, які змінюють інтенсивність виділення гормонів у відповідь на зміну концентрації певних речовин у внутрішньому середовищі організму.
Як уже вказувалося, нервова система, поряд із нервовими імпульсами, спроможна синтезувати і виділяти різноманітні хімічні речовини, які виконують функцію медіаторів і модифікаторів. Два відділи нервової системи – проміжний мозок, його гіпоталамічна частина, і ділянка спинного мозку спроможні виділяти речовини, що змінюють стан ендокринної системи. У гіпоталамусі виділяються рилізинг-фактори, що посилюють (ліберини) або послаблюють (статини) секрецію гормонів передньої частки гіпофіза.
У свою чергу гормони передньої частки гіпофіза можуть бути прямої дії (соматотропний, пролактин і меланоцитстимулювальний) а також кринотропними, які впливають на інші залози, змінюючи їх роботу.
Епіфіз
Епіфіз (пінеальний орган), верхній мозковий придаток, складається із заповнених колоїдом клітин – пінеоцитів, пронизаних множиною судинних і нервових стовбурів. Епіфіз у міног – це рудиментарне тім'яне око, що зберігає основні деталі будови цього органа. У хрящових і кісткових риб епіфіз звичайно добре розвинений, і має клітини, що нагадують клітини сітківки ока. У деяких риб епіфіз реагує на світло і бере участь у регуляції забарвлення. Для епіфіза кісткових риб характерна незакінченість трансформації із органа чуття в ендокринну залозу. Функції епіфіза зв'язують зі здійсненням циркадних ритмів діяльності організму і сезонної перебудови обміну речовин. Сигналом до такої перебудови слугує зміна довжини світлового дня.
Епіфіз є надзвичайно чутливим фотонейроендокринним перетворювачем сигналів зовнішнього середовища у гормональну відповідь. Навіть невеличкі зміни рівня освітленості викликають різкі зміни секреторної активності цієї фотосенсорної структури мозку хребетних. У епіфізі виявлені гормони серотонінімелатонін. Це низькомолекулярні речовини, які легко проникають крізь мембрани, є похідними амінокислоти триптофану. Серотонін викликає звуження дрібних артеріол, що веде до підвищення кров'яного тиску. Він же, крім того, слугує медіатором у деяких синапсах. Мелатонін, очевидно, є головним посередником, через який епіфіз впливає на ріст організму. Він гальмує розвиток статевих залоз, а також викликає контрактацію меланофорів і побіління шкіри. Відома імунорегулювальна дія гормону мелатоніну на організм. Наслідком гіпофункції епіфізу під час експерименту в результаті зниження секреції мелатоніну на світлі був зсув лейкоцитарної формули вбік росту кількості поліморфноядерних клітин і зниження числа лімфоцитів, наслідком гіперфункції в результаті підвищення секреції мелатоніну у темряві – підвищення виживаності (завдяки росту імунного статусу), зсув лейкоцитарної формули вбік зниження кількості поліморфноядерних клітин і росту кількості лімфоцитів, а також підвищення утворення в організмі інтерферону. Епіфізарна регуляція дозволяє ефективно боротися із грибковими захворюваннями за рахунок мобілізації імунітету організму.
Секреція мелатоніну в епіфізі – гормональна відповідь на отриману ним фотоінформацію, характеризується циклічністю: максимальна його кількість виділяється вночі, мінімальна – вдень.
Під час епіфізоектомії порушуються процеси синхронізації світлодинаміки і дозрівання гонад. Виявлено вплив епіфіза на ріст, а також на стан пігментних клітин. Штучна зміна річної світлодинаміки може вплинути також на статевий цикл – прискорити або відтягнути статеве дозрівання.
Гіпофіз
Гіпофіз розташований безпосередньо під гіпоталамусом на вузькій ніжці із нервової тканини і кровоносних судин. Він знаходиться посередині поглиблення в парасфеноїдній кістці.
У гіпофізі виявлено декілька гормонів. У нейрогіпофізі (частина гіпофіза, що ембріологічно утворюється із нервової тканини і пов'язана нервовими зв'язками із ядрами гіпоталамуса) накопичуються два гормони: окситоцин і аргінін – вазотоцин (АВТ), останній відрізняється від окситоцину лише заміною амінокислоти лейцину на аргінін.
У риб головна роль як окситоцину, так і АВТ, осморегулювальна. АВТ викликає підвищене утворення сечі у прісноводних риб, збільшуючи об'єм гломерулярної фільтрації. Окситоцин впливає на осморегуляцію, викликаючи звуження кровоносних судин у зябрах. АВТ викликає скорочення непосмугованих м'язів артеріол, що стабілізує або підвищує кров'яний тиск (що спостерігається у одних видів риб і відсутнє у інших) і може також впливати на осморегуляцію.
Із 7 гормонів, які виробляються передньою долею гіпофіза, три гормони безпосередньої дії, тобто вони діють на клітини або органи – мішені. Це пролактин, соматотропін і меланоцитстимулювальний гормон.
Пролактин (у риб його називають також паралактином) бере участь переважно в осморегуляції риб у прісній воді й у зв'язку із цим діє на багато органів. Він в основному сприяє утриманню деяких іонів епітелієм зябер, нирок і сечового міхура, а також зменшенню проникності зовнішніх покривів риб для води. Так, видалення гіпофіза у вугра прискорює втрату іонів через зябра, викликає загибель у деяких риб, пов'язану із втратою іонів.
Пролактин викликає також збільшення швидкості гломерулярної фільтрації у нирках. Він збільшує АТФ-азну активність у нирках і сечовому міхурі, але зменшує її у зябрах. Під дією пролактина зменшується поглинання води із кишечнику і сечового міхура, а також надходження води через зябра у результаті зниження проникності епітелію. Поглинання Na+ у нирках і сечовому міхурі зростає, у той час як виділення Na+ у зябрах зменшується. Кількість слизу, що утворюється у зябрах, кишечнику і шкірі, при цьому збільшується.
Соматотропін(СТГ) (який називають також гормоном росту) діє, як і пролактин, на багато тканин і органів без участі якоїсь проміжної ендокринної залози. Цей гормон в основному стимулює апетит і ріст риб, а також запобігає гіпертрофії печінки. Бичачий гормон росту в разі внутрішньом’язового введення один раз у тиждень прискорює ріст кижуча на 50 – 100 % порівняно із контролем.
Меланоцитстимулювальний гормон(МСГ) надходить із гіпофіза і викликає експансію меланоцитів у шкірі риб, у результаті чого риби темнішають. Але МСГ діє не на всі пігментні клітини. Існує декілька типів пігментних клітин (хроматофорів), кожний із яких регулюється самостійно. Крім того, хроматофори одного типу можуть по-різному реагувати на однаковий стимул залежно від місцезнаходження.
Крім гормонів безпосередньої дії гіпофіз виділяє низку кринотропних гормонів, тобто гормонів, що посилюють активність інших ендокринних залоз. До таких гормонів відносяться: адренокортикотропний (АКТГ), тиреотропний (ТТГ) і гонадотропіни. АКТГ стимулює утворення кортизолу та інших кортикостероїдів інтерреналовими клітинами. ТТГ стимулює утворення у щитоподібній залозі або звільнення із неї тироксину. Гонадотропіни (у деяких риб, можливо, тільки один гонадотропін), стимулюють утворення гамет у гонадах костистих риб.
Н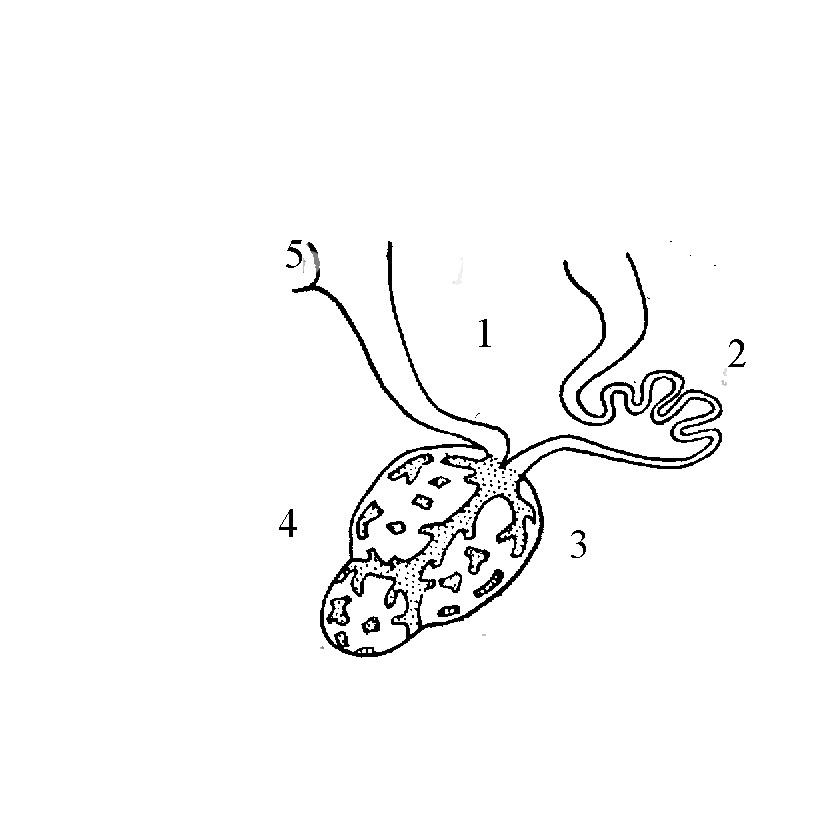 айменшій
вміст гонадотропінів у гіпофізі
спостерігається після нересту, а
найбільший – перед нерестом. Вивчення
вмісту кожного із гонадотропінів у
гіпофізі коропа і білого товстолобика
показало, що їх динаміка різна і, можливо,
вони відіграють у статевому процесі
риб приблизно ту ж роль, що і
фолікулостимулювальний (ФСГ) та
лютенізуючий (ЛГ) гормони у ссавців,
тобто, інтенсивність росту овоцитів
регулюється гормоном, подібним ФСГ, а
реакція дозрівання пов'язана із гормоном,
близьким до ЛГ.
айменшій
вміст гонадотропінів у гіпофізі
спостерігається після нересту, а
найбільший – перед нерестом. Вивчення
вмісту кожного із гонадотропінів у
гіпофізі коропа і білого товстолобика
показало, що їх динаміка різна і, можливо,
вони відіграють у статевому процесі
риб приблизно ту ж роль, що і
фолікулостимулювальний (ФСГ) та
лютенізуючий (ЛГ) гормони у ссавців,
тобто, інтенсивність росту овоцитів
регулюється гормоном, подібним ФСГ, а
реакція дозрівання пов'язана із гормоном,
близьким до ЛГ.

