

Ефективність поглинання кисню та енергетичні витрати на дихання
Оскільки дихальні насоси досить універсальні щодо активності риб і вмісту кисню у воді, важко зробити широкі узагальнення, що стосуються їхніх можливостей і обмежень. Загальним для всіх їх є те, що ефективність поглинання кисню (відсоток доступного кисню, що поглинається із води у зябрах) знижується під час зростання вентиляційного об'єму. Чим менша активність риби і вентиляційний об'єм, тим більший коефіцієнт поглинання кисню із води. Виявлено, що така донна малорухома риба, як морський язик, поглинає у стані спокою до 80 % доступного О2. Необхідною умовою для цього є пристасованість більшості тканин до дуже низького вмісту кисню, що припускає низьке венозне Po2. Якби Po2 води був більш низьким, кисень дифундував би із крові у воду.
Ефективність поглинання кисню (або утилізація) коливається і залежить від активності риби. У лососевих ефективність поглинання кисню із води із посиленням вентиляції зябер дещо знижується. У золотої рибки і коропа ефективність поглинання кисню у стані спокою висока, а у активному стані – дуже низька. Відповідно до вентиляційних об'ємів і ефективності поглинання кисню енергія, що витрачається, також коливається у широкому діапазоні. У лососевих риб енергетичні витрати на дихання еквівалентні 0,5 % споживаного кисню у стані спокою і до 15 % в активному стані. Це означає, що за умов збільшення споживання кисню в 5 разів фактичні енергетичні витрати зростають приблизно в 150 разів. У лина енергетичні витрати на дихальну вентиляцію змінюються від 18 – 43 % споживаного кисню у стані спокою до 44 – 69 % при збільшенні вентиляційного об'єму у 3 рази. Якщо це порівняти із енергетичними витратами у стані спокою людини (біля 2 % споживаного кисню) стає ясно, що риби дійсно витрачають більше енергії на прокачування через зябра води замість повітря. Необхідно також відзначити, що у риб, що дихають повітрям, площа зябрової поверхні у 5 разів менша, ніж у близькородинних риб, що ведуть цілком водний спосіб життя, тобто водне дихання дійсно потребує розвинених дихальних поверхонь для зведення до мінімуму енергетичних витрат на вентиляцію зябер.
Характеристика протиточної обмінної системи
У зв'язку із тим, що транспорт кисню із води до крові відбувається тільки шляхом дифузії, за багато мільйонів років природного добору у риб виробився високоефективний механізм інтенсифікації дифузії кисню. Таким механізмом є протиточна обмінна система (мал. 11.1).
Протиточні обмінні системи більш ефективні у порівнянні із системами, у яких потоки рідин в обох її частинах мають однаковий напрямок. У системі із однонаправленими потоками градієнт дифузії спочатку високий і кількість поглинутого О2 також значна, але потім швидко знижується, поки у двох потоках не досягне рівноваги. В цей час із одного потоку в інший не може бути перенесена більш як половина розчиненого кисню. Зміна напрямку руху одного із потоків призводить до утворення цілком нових умов. Венозна кров, отримавши деяку кількість кисню через мембрани із води, що протікає, далі рухається в умовах, за яких вміст кисню у воді, що притікає, постійно збільшується. Хоча деякі риби спроможні зв'язати 80 % розчиненого кисню, можливість поглинання більш половини доступного кисню цілком реальна завдяки майже постійному градієнту дифузії всією довжиною протиточної системи.
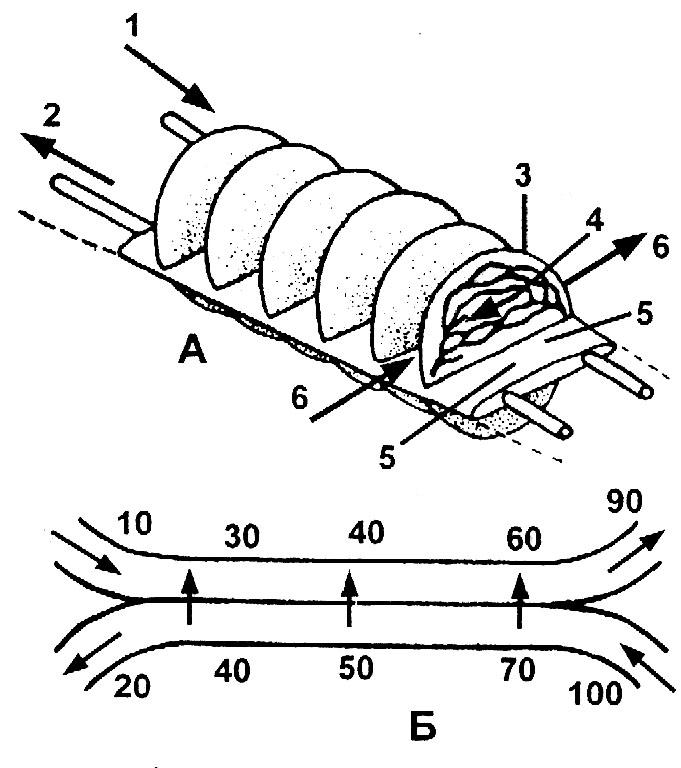
Рис. 11.2. Схема протиточної системи в зябрах риб:А – фрагмент зябрового пелюстка с зябровими пластинками: 1 – артерія; 2 – вена; 3 – зяброва пластинка; 4 – кров; 5 –зябровий пелюсток; 6 – вода; Б – принципова схема протиточного обмінника (цифрами позначено ступінь насиченості газами води)
Протиточні обмінні системи більш ефективні у порівнянні із системами, у яких потоки рідин в обох її частинах мають однаковий напрямок. У системі із однонаправленими потоками градієнт дифузії спочатку високий і кількість поглинутого О2 також значна, але потім швидко знижується, поки у двох потоках не досягне рівноваги. В цей час із одного потоку в інший не може бути перенесена більш як половина розчиненого кисню. Зміна напрямку руху одного із потоків призводить до утворення цілком нових умов. Венозна кров, отримавши деяку кількість кисню через мембрани із води, що протікає, далі рухається в умовах, за яких вміст кисню у воді, що притікає, постійно збільшується. Хоча деякі риби спроможні зв'язати 80 % розчиненого кисню, можливість поглинання більш половини доступного кисню цілком реальна завдяки майже постійному градієнту дифузії всією довжиною протиточної системи.
Протитоки спостерігаються тільки на найтоншому рівні організації зябер – у зябрових пластинках, що неозброєним оком не помітні, за винятком тільки найбільших риб. Пластинки виступають із дорсальної і вентральної поверхонь кожного зябрового пелюстка і можуть розташовуватися між пластинками суміжного зябрового пелюстка. Кожна пара цих пелюстків розташована на зябрових дугах V-подібно, таким чином, що вода рухається тільки через ці структури, а не навколо них. Кінці зябрових пелюстків суміжних зябрових дуг часто стикаються, тому вода через зябра може проходити тільки між пелюстками. Хоча зяброві пелюстки у більшості видів риб посилені хрящовим тяжем, V-подібна форма може порушуватися під дією швидкісного руху води, наприклад під час швидкого плавання. При цьому значна частина води може проходити повз пелюстки і газообмін не відбувається. У цьому полягає одна із причин зниження ефективності поглинання кисню із води під час високих швидкостей вентиляції. У деяких риб, що швидко плавають, наприклад, тунців, кінці двох суміжних зябрових пелюстків сполучені, що запобігає порушенню V-подібної форми зябрових пелюстків і підтримує максимальний газообмін при високих швидкостях руху води.
Коли вода постійно протікає повз пластинки в одному напрямку, протиточний обмін легко здійснюється, оскільки у пластинках кров рухається у зворотному напрямку. Зяброві приносні артерії на внутрішньому боці зябрових пластинок і зяброві виносні артерії на зовнішньому, омиваному водою боці утворюють у зябрових пластинках протиток.
Кров рухається у тонкому шарі пластинок, товщина яких підтримується опорними клітинами, а відстань дифузії мала, що полегшує поглинання кисню. Товщина тканини пластинок, через яку кисень переходить із води до крові, складає 1 – 5 мкм.