

Слухова сенсорна система
Рецепція звуку може здійснюватися в декількох відділах внутрішнього вуха риб: сакулюсі, утрикулюсі і лагені. Хоча шляхи збудження цих структур можуть бути різними, в усіх випадках безпосереднім рецептором механічних коливань також є вторинночутливі волоскові клітини. У найпростішому випадку звук може викликати збудження рецептора за рахунок різної інерційності утворень, пов'язаних із утвореннями волоскової клітини і із вершиною її волосків. Такий сигнал призведе до зсуву волосків, що викликає появу внутрішньоклітинного рецепторного потенціалу (мал. 4.3).
У циприноїдних риб основним приймачем звуку є плавальний міхур, коливання якого через спеціалізований апарат кісточок Вебера передаються у рідкі середовища внутрішнього вуха.
За обох зазначених засобах збудження рецепторів ефективність сигналу буде залежати від напрямку його приходу. Асиметрія приймальних експериментах, які показали, що найбільша чутливість спостерігається при за надходження звуку із переднього напрямку.
Асиметрія приймачів внутрішнього вуха обумовлена просторовою організацією отолітів і рецепторних клітин. Деполяризація рецепторів відбувається під час зсуву пучка жорстких волосків (стереоцилій) у напрямку до одного гнучкого волоска – кіноцилії. У кожному із згаданих слухових відділів внутрішнього вуха риб є видоспецифічна топологічна організація клітин, що мають по-різному орієнтовані пучки волокон. Ця топографія супроводжується відповідною організацією гілочок слухового нерва. У результаті варто очікувати спеціалізації гілочок за чутливістю до напрямку приходу звуку. У циприноїдних риб сакулярна макула витягнута горизонтально. У горизонтальній частині макули кіноцилії розташовані, як правило, із дорсальної сторони пучка стереоцилій, а у вертикальній частині – із вентральної. Сакулярний отоліт цих риб порівняно легкий, і його функціонально можна зіставити із текторіальною мембраною наземних хребетних. Вигин стереоцилій викликається рухом ендолімфи, що призводить до коливань отоліту.
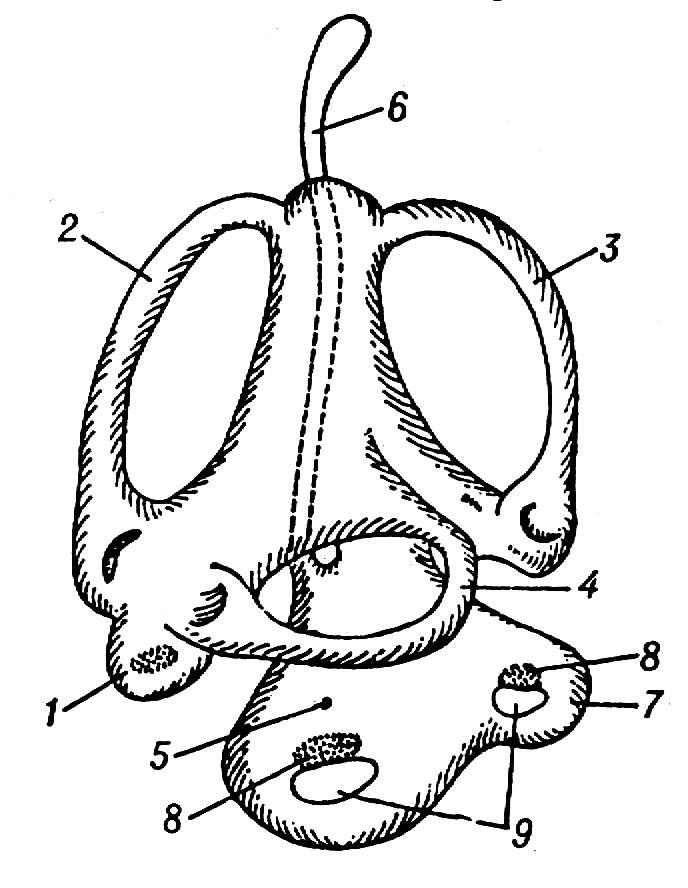
Мал. 4.3. Внутрішнє вухо риб:1 – овальний мішечок (утрикулюс); 2 – 4 – півколові канали; 5 – круглий мішечок (сакулюс); 6 – ендолімфатичний канал; 7 – завитка (лагена); 8 – слухові чутливі поля; 9 – отоліти
У більшості інших риб, що не мають спеціальної системи передачі звуків до внутрішнього вуха від плавального міхура, сакулярний отоліт звичайно більш масивний. В зв’язку із тим, що щільність отоліту приблизно у три рази перевищує щільність води і макулярних клітин, його зсуви запізнюються щодо тіла клітини. Результуючий вигин волосків обумовлює деполяризацію волоскової клітини і збудження дендритів нейронів, аксони яких формують слуховий нерв. Топографія клітин із різною орієнтацією волосків дуже варіабільна і може бути цілком різною навіть у видів, що належать до одного сімейства.
Порівняно легкий отоліт циприноїдних може коливатися у різних точках по-різному в залежності від частоти. У риб із масивним отолітом характер його зсуву стосовно більш рухливої макули також змінюється при зміні частоти звуку. У результаті ділянка збудження рецепторів у обох групах риб є частотнозалежною.
Від слухових утворень внутрішнього вуха йдуть тонкі гілочки, що, зливаючись із гілочками від інших рецепторних структур, утворюють передній і задній рукав 8-го черепного нерва. У сріблястого карася волокна сакулярної гілочки варіюють за діаметром від 2 до 10 мкм, причому одне волокно звичайно пов'язане із декількома рецепторними клітинами.
Слухове волокно 8-го нерва закінчуються в декількох ядрах довгастого мозку, номенклатура яких дотепер установлена не зовсім чітко і, мабуть, істотно різниться у різних груп. Більшість волокон розгалужуються, посилаючи один паросток у передню групу ядер, а інший у задню. Відростки, що йдуть уперед, можуть досягати вентролатеральної частки мозочка, а ті, що відходять назад – маунтеровських клітин. Нещодавно в довгастому мозку риб описане вторинне слухове ядро, входом якого слугують аксони нервових клітин другого порядку.
Слухові нейрони довгастого мозку посилають аксони у середній мозок, закінчуючись майже винятково в медіальних відділах напівколового торуса. Проекції у проміжний і передній мозок дотепер фактично не досліджені.
Риби спроможні сприймати дуже різноманітні звукові сигнали. Звуки небіологічного походження, що виникають, наприклад, внаслідок руху води, можуть використовуватися для оцінки метеоумов або визначення гідрологічних змін. Безсумнівно, що риби сприймають звуки суден і інших штучних об'єктів.
Більш ретельні дослідження слухових спроможностей риб зроблені в лабораторних умовах із використанням синтезованих сигналів. Критерієм наявності сприйняття використовували як безумовні, так і умовні рефлекторні реакції. У більшості випадків частотний оптимум відповідає сотням герц при щонайнижчих порогах 20 – 50 децибел рівня звукового тиску. Верхня межа аудіограм варіює від 0,3 до 3,0 кГц. Але дотепер немає повної впевненості в тому, що ці розходження не пов'язані із методикою експериментальних робіт, проведених різними групами авторів. Проте варто відзначити, що найбільш високі пороги і найменший частотний діапазон сприйняття має риба-жаба, що відрізняється особливо голосним звуковипромінюванням. У сріблястого карася (Carassius auratus) дуже швидко виробляється умовний рефлекс, що полягає в гальмуванні дихальних рухів після впливу звуку. Ця методика дозволила одержати не тільки абсолютні пороги відчуття, але також диференціальні пороги і пороги маскування. Діапазон відчуття карася простирається до 2,0 кГц із широким оптимумом аудіограми у межах 0,3 – 0,9 кГц. Для визначення ступеня частотного дозволу в цього об'єкта використовували метод маскування тоном. Поріг маскування був найбільшим за збігом частот тестового тону із маскера і досить різко падав при зміні будь-якої із цих частот. Результати із безсумнівністю свідчать про можливість частотного аналізу звуків. Диференціальна чутливість до частоти тонів, обмірювана тим же методом, складала біля 5 %.
Однією із найбільш принципових проблем, що стосуються слуху риб, уже давно було питання про спроможність цих тварин до локалізації джерела звуку. Поведінкові експерименти, проведені в умовах відкритих водних басейнів при розташуванні джерела в дальньому звуковому полі, де бічна лінія не функціонує, ясно демонструє непогані спроможності до локалізації як у хрящових так і костистих риб. Більш того, у тріски виявлена досить висока диференціальна чутливість за кутом не тільки в горизонтальній, але й у вертикальній площині. Точність локалізації сигналу по вертикалі складає 10 – 15 %, що перевищує спроможності більшості ссавців, у тому числі і людини. Функціональне значення цієї особливості риб очевидно, тому що водний простір принципово трьохмірний.
Більшість риб сприймає звук у діапазоні від 16 Гц до 4 – 5 кГц (деякі види до 13 кГц). Найменша інтенсивність звуку, що сприймають риби, варіює в межах від 0 до 70 децибел. У риб, що мають плавальний міхур, відчуття звуку більш виражене. Плавальний міхур при цьому відіграє роль перетворювача звукових хвиль у механічні коливання, які сприймаються слуховими рецепторами. Велике значення при цьому має веберів апарат. У безміхурних риб подібну трансформацію здійснюють повітряні порожнини, пов'язані із внутрішнім вухом.
За гостротою слуху декотрі риби не поступаються людині, розрізняючи 1/4 тону – 1/2 октави. Пропускна спроможність слухового апарата така ж, як і зору, а перешкодостійкість (відношення сигнал/шум) приблизно 1:1. Сприйняття акустичних коливань у риб – спрямоване. Але акустична база (відстань між рецепторами) у риб невелика – менше 1 см, звідси й обмеженість бінаурального ефекту, тобто спроможності визначати положення джерела звуку за різницею приходу звукових коливань на обидва рецептори. Звичайно дальність орієнтування на джерело звуку за допомогою слухових рецепторів обмежується декількома метрами. Електрофізіологічні дослідження слухової системи риб розвиваються вкрай нерівномірно. Є серйозні роботи, які стосуються імпульсної активності волокон слухового нерва, але характеристики нейронів центральних утворень слухового шляху вивчені явно недостатньо.
У слухових центрах довгастого мозку тріски нейрони мають спонтанну активність, а реакція на тони є частотно вибірковою. Проте на відміну від слухового нерва синхронізація із тоном у більшості випадків виявляється досить слабкою. Відповідь на звук спостерігається і у маунтеровських клітинах, приводячи до скорочення м'язів контрлатеральної сторони тіла, що обумовлюють так називаний старт-рефлекс.
У торусі Ictalurus punctatus на звук відповідає приблизно половина клітин, причому реакції могли бути як збудливими, так і гальмівними. Використовуючи метод електричної стимуляції, удалося показати, що деякі клітини торусу одержують білатеральні входи.
У деяких точках вентролатеральної зони таламуса хрящових риб спостерігали викликану відповідь на звукові стимули. Відповідь не викликалася стимуляцією електрорецепторів і органів бічної лінії.
Ці досить уривчасті дані хоча і не дозволяють виявити послідовні етапи опрацювання сигналу, але в цілому укладаються в загальну схему ієрархічної організації ознакового опису сигналу в слуховому шляху. Зазначимо основні риси цієї схеми: максимально повне відтворення сигналу у волокнах слухового нерва, виділення частотно-часових особливостей і первинне бінауральне порівняння в ядрах довгастого мозку, виділення більш складних ознак у середньому мозку. Особливістю слухової системи риб є порівняно вузький частотний діапазон відтворених звуків і домінування часових механізмів аналізу акустичних сигналів.