- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с. Предисловие редакторов перевода
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с. Предисловие автора к русскому изданию
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с. Предисловие
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с. Часть I. Как происходит развитие Глава 1. Введение в изучение развития животных
- •Введение
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •12___________________ Глава 1________________________________________________________________________________________________
- •Основные особенности развития
- •Наше эукариотическое наследие
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •16_______________ Глава 1________________________________________________________________________________
- •Развитие одноклеточных эукариот
- •Регуляция морфогенеза при развитии ацетабулярии
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дифференцировка у амебофлагеллятыNaegleria
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Происхождение полового размножения
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Колониальные эукариоты: эволюция дифференцировки
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Пол и смерть уVolvocales
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Слизевик Dictyostelium
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •30________________ Глава 1_______________________________________________________________________________
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Особенности развития разных многоклеточных
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •32________________ Глава 1________________________________________________________________________________
- •Первичноротые и вторичноротые
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Цитированная литература
- •Строение гамет
- •Спермий
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •36________________ Глава 2________________________________________________________________________
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •38________________ Глава 2___________________________
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •40________________ Глава 2______________________________________________________________________________
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Узнавание спермия и яйца: воздействие на расстоянии
- •Привлечение спермиев
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Акросомная реакция
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Узнавание спермия и яйца: контакт между гаметами Видоспецифическое узнавание у морских ежей
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •46 Глава 2
- •Соединение гамет у млекопитающих
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •50________________ Глава 2______________________________________________________________________________
- •Слияние гамет и предотвращение полиспермии Слияние плазматических мембран спермия и ооцита
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Предотвращение полиспермии
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •56________________ Глава 2______________________________________________________________________________
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Слияние генетического материала
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Неравнозначность пронуклеусов у млекопитающих
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Активация метаболизма яйца
- •Ранние реакции
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Поздние реакции
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Биохимия активации яйца
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Преобразования цитоплазмы яйца
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Цитированная литература
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Пространственные закономерности дробления зародыша
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Радиальное голобластическое дробление Морской огурецSynapta
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Морские ежи
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Амфибии
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Спиральное голобластическое дробление
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Адаптация путем изменения процесса дробления зародыша
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Билатеральное голобластическое дробление
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Чередующееся (rotational) голобластическое дробление
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Компактизация
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •90 Глава 3
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Клеточная поверхность и механизм компактизации
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Образование внутренней клеточной массы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Освобождение от прозрачной оболочки
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Близнецы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Меробластические типы дробления
- •Дискоидальное дробление
- •Поверхностное дробление
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Механизмы дробления Инициация дробления
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •102 Глава 3
- •Цитоскелетный механизм митоза
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Образование новых мембран
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Цитированная литература
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гаструляция у морского ежа
- •Выселение первичной мезенхимы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Первая фаза инвагинации
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Вторая фаза инвагинации
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гаструляция у амфибий
- •Движения клеток в период гаструляции у амфибий: обзор
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Детерминация положения бластопора
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Клеточные движения и образование архентерона
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Миграция инволюирующей мезодермы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •128 Глава 4
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Эпиболия эктодермы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •130 Глава 4
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гаструляция у птиц Основные черты гаструляции у птиц
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Механизмы гаструляции у птиц
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •136 Глава 4
- •Гаструляция у млекопитающих
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •138________________ Глава 4
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •140 Глава 4
- •Дополнительные сведения и гипотезы: Функция хориона
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •142 Глава 4
- •Цитированная литература
- •Основные черты развития позвоночных
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Нейруляция
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Нервная трубка и происхождение центральной нервной системы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Механизм формирования нервной трубки
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дифференцировка нервной трубки
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Эволюция развития головного мозга
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Развитие глаза
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дифференцировка хрусталика и роговицы.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Нервный гребень и его производные
- •Пути миграции клеток нервного гребня
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Полипотентность клеток нервного гребня
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Эпидермис и его производные
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •178_______________ Глава 5_______________________________________________________________________________
- •Дополнительные сведения и гипотезы: Тератология
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Цитированная литература
- •Дорсальная мезодерма: дифференцировка сомитов
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Миогенез: дифференцировка скелетной мышцы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Остеогенез: развитие кости
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Мезодерма боковых пластинок
- •Дополнительные сведения и гипотезы: Изоформы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Формирование внезародышевых оболочек
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Сердце и системы кровообращения
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Формирование кровеносных сосудов
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Изменение направления потока крови у новорожденного млекопитающего
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Ангиогенез, индуцированный опухолью
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Развитие клеток крови Полипотентные стволовые клетки и их микроокружение
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Локализация гемопоэза (кроветворения)
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Энтодерма
- •Пищеварительная трубка и ее производные
- •Печень, поджелудочная железа и желчный пузырь
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дыхательная трубка
- •Дополнительные сведения и гипотезы: Тератокарцинома
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с. Оглавление
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Глава 6. Раннее развитие позвоночных.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Часть I. Как происходит развитие 11
- •Глава 1. Введение в изучение развития животных 11
- •Глава 2 Оплодотворение: возникновение нового организма 35
- •Глава 3 Дробление: создание многоклеточности 74
- •Глава 4 Гаструляция: перераспределение клеток 110
- •Глава 5 Раннее развитие позвоночных. Производные эктодермы, их органо- и гистогенез 145
- •Глава 6 Раннее развитие позвоночных. Производные мезодермы и энтодермы, их органо- и гистогенез 186
Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
|
|
|
Рис. 1.3. Схематическое изображение митоза в животных клетках. Во время интерфазы при подготовке клетки к делению происходит репликация ДНК. Во время профазы ядерная оболочка разрушается и между двумя центриолями формируется веретено. На стадии метафазы хромосомы располагаются в экваториальной плоскости клетки. Когда наступает анафаза, удвоившиеся хромосомы (называемые хроматидами) расходятся. На стадии телофазы хромосомы достигают полюсов веретена, клетка начинает разделяться на две дочерние клетки. По числу и типу хромосом дочерние клетки идентичны материнской. |

|
|
|
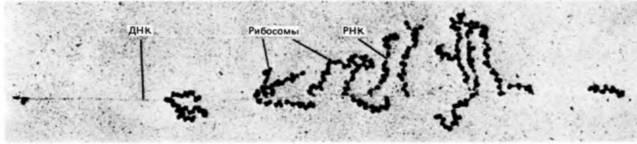
Рис. 1.4. Одновременность транскрипции и трансляции у прокариот. Участок ДНК Escherichia coli пересекает электронную микрофотографию по горизонтали. Транскрипты мРНК видны по обе стороны от нити ДНК. Рибосомы прикрепились к мРНК и синтезируют белки (которые не видны). Следует отметить, что длина синтезируемых мРНК увеличивается слева направо: это указывает на направление транскрипции. (С любезного разрешения О. L. Miller. Jr.) |
Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
16_______________ Глава 1________________________________________________________________________________
полимеразами. У прокариот новосинтезированная мРНК немедленно начинает транслироваться с образованием белка, в то время как другой конец молекулы еще продолжает транскрибироваться с ДНК (рис. 1.4). Таким образом, транскрипция и трансляция у прокариот осуществляются одновременно и координированно. Существование ядерной оболочки у эукариот делает возможным совершенно новый тип клеточной регуляции. Рибосомы, ответственные за трансляцию, находятся по одну сторону ядерной оболочки, тогда как ДНК, а также РНК-полимераза, необходимые для трансляции, по другую ее сторону. В период между транскрипцией и трансляцией транскрибированная РНК должна претерпеть такие изменения, которые позволили бы ей пройти через ядерную оболочку. Регулируя выход мРНК в цитоплазму, клетка способна отобрать те из вновь синтезированных носителей информации, которые должны транслироваться. Таким образом, сложность организации достигает нового уровня, который, как мы увидим, чрезвычайно важен для развивающегося организма.
Развитие одноклеточных эукариот
Все многоклеточные эукариотические организмы произошли в процессе эволюции от одноклеточных простейших. У этих простейших впервые появились основные особенности, присущие развитию эукариот. Примитивные эукариоты дают первые примеры морфогенеза, контролируемого ядром, использования клеточной поверхности как посредника для кооперации отдельных клеток, появления полового размножения.
Регуляция морфогенеза при развитии ацетабулярии
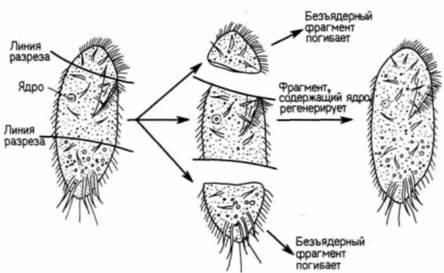
В конце XIX в. еще не было доказано, что в ядре содержится информация, определяющая наследственность и направляющая развитие. Одним из лучших свидетельств в пользу такого представления были результаты опытов по разделению одноклеточных организмов на фрагменты, содержащие ядро, и безъядерные (Wilson. 1896). Когда различных простейших разрезали на множество частей, почти все эти части погибали. Однако фрагменты, содержащие ядро, оказывались жизнеспособными и регенерировали все сложные клеточные структуры (рис. 1.5).
Ядерный контроль клеточного морфогенеза и взаимодействие между ядром и цитоплазмой прекрасно иллюстрируют опыты на ацетабулярии (Acetabularia). Эта гигантская одиночная клетка (длиной в 2.5 - 5 см) состоит из трех частей: шляпки, стебелька и ризоида (рис. 1.6). Ризоид находится в основании клетки и прикрепляет ее к субстрату. Единственное клеточное ядро располагается в ризоиде. Большие размеры ацетабулярии и положение ее ядра позволяют исследователям удалить из клетки ядро и заменить его ядром другой клетки. В 30-х годах нашего века Геммерлинг (J. Hämmerling) воспользовался этими уникальными свойствами ацетабулярии и пересадил ядро одного вида (Acetabularia mediterranea) в энуклеированный ризоид другого (A. crenulata). Как видно из рис. 1.6. шляпки у этих видов очень сильно отличаются друг от друга. Геммерлинг обнаружил, что после того, как ядро одного вида трансплантировали в стебелек другого, новообразующаяся шляпка приобретала строение, свойственное
|
|
Рис. 1.5. Регенерация содержащего ядро фрагмента одноклеточного простейшего Stylonychia. Безъядерные фрагменты сохраняются в течение некоторого времени, но в конце концов погибают. |