

- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с. Предисловие редакторов перевода
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с. Предисловие автора к русскому изданию
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с. Предисловие
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с. Часть I. Как происходит развитие Глава 1. Введение в изучение развития животных
- •Введение
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •12___________________ Глава 1________________________________________________________________________________________________
- •Основные особенности развития
- •Наше эукариотическое наследие
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •16_______________ Глава 1________________________________________________________________________________
- •Развитие одноклеточных эукариот
- •Регуляция морфогенеза при развитии ацетабулярии
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дифференцировка у амебофлагеллятыNaegleria
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Происхождение полового размножения
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Колониальные эукариоты: эволюция дифференцировки
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Пол и смерть уVolvocales
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Слизевик Dictyostelium
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •30________________ Глава 1_______________________________________________________________________________
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Особенности развития разных многоклеточных
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •32________________ Глава 1________________________________________________________________________________
- •Первичноротые и вторичноротые
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Цитированная литература
- •Строение гамет
- •Спермий
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •36________________ Глава 2________________________________________________________________________
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •38________________ Глава 2___________________________
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •40________________ Глава 2______________________________________________________________________________
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Узнавание спермия и яйца: воздействие на расстоянии
- •Привлечение спермиев
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Акросомная реакция
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Узнавание спермия и яйца: контакт между гаметами Видоспецифическое узнавание у морских ежей
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •46 Глава 2
- •Соединение гамет у млекопитающих
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •50________________ Глава 2______________________________________________________________________________
- •Слияние гамет и предотвращение полиспермии Слияние плазматических мембран спермия и ооцита
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Предотвращение полиспермии
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •56________________ Глава 2______________________________________________________________________________
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Слияние генетического материала
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Неравнозначность пронуклеусов у млекопитающих
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Активация метаболизма яйца
- •Ранние реакции
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Поздние реакции
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Биохимия активации яйца
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Преобразования цитоплазмы яйца
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Цитированная литература
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Пространственные закономерности дробления зародыша
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Радиальное голобластическое дробление Морской огурецSynapta
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Морские ежи
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Амфибии
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Спиральное голобластическое дробление
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Адаптация путем изменения процесса дробления зародыша
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Билатеральное голобластическое дробление
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Чередующееся (rotational) голобластическое дробление
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Компактизация
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •90 Глава 3
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Клеточная поверхность и механизм компактизации
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Образование внутренней клеточной массы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Освобождение от прозрачной оболочки
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Близнецы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Меробластические типы дробления
- •Дискоидальное дробление
- •Поверхностное дробление
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Механизмы дробления Инициация дробления
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •102 Глава 3
- •Цитоскелетный механизм митоза
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Образование новых мембран
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Цитированная литература
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гаструляция у морского ежа
- •Выселение первичной мезенхимы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Первая фаза инвагинации
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Вторая фаза инвагинации
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гаструляция у амфибий
- •Движения клеток в период гаструляции у амфибий: обзор
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Детерминация положения бластопора
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Клеточные движения и образование архентерона
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Миграция инволюирующей мезодермы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •128 Глава 4
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Эпиболия эктодермы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •130 Глава 4
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гаструляция у птиц Основные черты гаструляции у птиц
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Механизмы гаструляции у птиц
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •136 Глава 4
- •Гаструляция у млекопитающих
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •138________________ Глава 4
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •140 Глава 4
- •Дополнительные сведения и гипотезы: Функция хориона
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •142 Глава 4
- •Цитированная литература
- •Основные черты развития позвоночных
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Нейруляция
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Нервная трубка и происхождение центральной нервной системы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Механизм формирования нервной трубки
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дифференцировка нервной трубки
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Эволюция развития головного мозга
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Развитие глаза
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дифференцировка хрусталика и роговицы.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Нервный гребень и его производные
- •Пути миграции клеток нервного гребня
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Полипотентность клеток нервного гребня
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Эпидермис и его производные
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •178_______________ Глава 5_______________________________________________________________________________
- •Дополнительные сведения и гипотезы: Тератология
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Цитированная литература
- •Дорсальная мезодерма: дифференцировка сомитов
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Миогенез: дифференцировка скелетной мышцы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Остеогенез: развитие кости
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Мезодерма боковых пластинок
- •Дополнительные сведения и гипотезы: Изоформы
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Формирование внезародышевых оболочек
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Сердце и системы кровообращения
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Формирование кровеносных сосудов
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Изменение направления потока крови у новорожденного млекопитающего
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дополнительные сведения и гипотезы: Ангиогенез, индуцированный опухолью
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Развитие клеток крови Полипотентные стволовые клетки и их микроокружение
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Локализация гемопоэза (кроветворения)
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Энтодерма
- •Пищеварительная трубка и ее производные
- •Печень, поджелудочная железа и желчный пузырь
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Дыхательная трубка
- •Дополнительные сведения и гипотезы: Тератокарцинома
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с. Оглавление
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Глава 6. Раннее развитие позвоночных.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
- •Часть I. Как происходит развитие 11
- •Глава 1. Введение в изучение развития животных 11
- •Глава 2 Оплодотворение: возникновение нового организма 35
- •Глава 3 Дробление: создание многоклеточности 74
- •Глава 4 Гаструляция: перераспределение клеток 110
- •Глава 5 Раннее развитие позвоночных. Производные эктодермы, их органо- и гистогенез 145
- •Глава 6 Раннее развитие позвоночных. Производные мезодермы и энтодермы, их органо- и гистогенез 186
Миграция инволюирующей мезодермы
Миграция клеток-предшественников мезодермы внутрь зародыша является частью удивительно хорошо координированной серии глобальных клеточных миграций. Подробная схема этих движений приведена на рис. 4.25. Один из наиболее важных аспектов гаструляции у амфибий это схождение к центру (конвергенция) и удлинение (растяжение) инволюирующей краевой зоны (ИКЗ), которая представляет собой область, расположенную непосредственно над губой бластопора. В поверхностном слое этой зоны (ИКЗП0В) находится материал энтодермальной крыши архентерона, а ее глубокий слой (ИКЗГЛ) содержит проспективные мезодермальные клетки (в том числе клетки хорды).
Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
_______________ ГАСТРУЛЯЦИЯ ПЕРЕРАСПРЕДЕЛЕНИЕ КЛЕТОК 127
|
|
|
Рис. 4.24. Интегрированная модель клеточных движений в период гаструляции. А. Ранняя гаструла характеризуется интердигитацией слоев глубоких клеток краевой зоны и их инволюцией. Б. В. Зародыши на более поздних стадиях гаструляции. Глубокие краевые клетки уплощаются, а бывшие поверхностные клетки краевой зоны образуют стенку архентерона (гастроцеля). Колбовидные клетки выделены частыми точками. (По Keller. 1981.) |
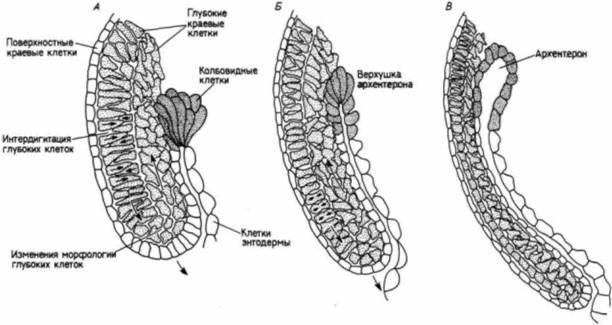
Во время гаструляции клетки анимальной чаши и неинволюирующей краевой зоны распространяются путем эпиболии, покрывая весь зародыш. Дорсальная часть неинволюировавших краевых клеток расширяется быстрее, чем вентральная, заставляя губы бластопора двигаться в вентральном направлении. Клетки из этих двух зон будут формировать эктодерму зародыша. Глубокая инволюирующая краевая зона представляет собой кольцо клеток, из которых образуются хорда, головная мезодерма, мезодерма сомитов и боковых пластинок. Во время гаструляции это кольцо инволюирует через губы бластопора, формируя мезодермальную мантию. Клетки-предшественники головной мезодермы, боковой и брюшной мезодермы распространяются во всю ширину зародыша, тогда как материал хорды и сомитов конвергирует в дорсальном направлении, образуя дорсальные осевые структуры. Клетки поверхностной инволюирующей краевой зоны также претерпевают конвергентное растяжение формируя энтодермальные ткани крыши архентерона. Суббластопоральная энтодерма покрывается обрастающими ее клетками неинволюирующей краевой зоны и образует дно архентерона (Keller, I986).
В процессе конвергентного растяжения ИКЗГЛ ее клетки сами перестраиваются, вставляясь между прилежащими к ним клетками и создавая тем самым тонкое удлиненное образование вместо первоначально широкой маленькой полоски клеток (рис. 4.26). Мы уже обсуждали сходный феномен при описании гаструляции у морского ежа, включающий в себя растяжение архентерона. Из многочисленных рядов клеток путем интеркаляции образуется меньшее число более длинных рядов клеток. Конвергентное растяжение мезодермы является, по-видимому, автономным, потому что движения клеток наблюдаются даже в том случае, когда они изолированы из остальной части зародыша (Keller, 1986).
Как инволюирующие клетки узнают, куда им надо двигаться сразу же после попадания внутрь зародыша? Имеются ли на внутренней поверхности крыши бластоцеля некие молекулы, облегчающие миграцию клеток? По-видимому, инволюирующие клетки-предшественники мезодермы мигрируют к анимальному полюсу по фибронектиновой сети, секретируемой клетками крыши бластоцеля. У амфибий незадолго до начала гаструляции презумптивная эктодерма крыши бластоцеля секретирует внеклеточный матрикс, который содержит волокна фибронектина (Boucaut et al., 1984; Nakatsuji et al., 1985) (рис. 4.27). Подобно инвагинирующей мезенхиме морского ежа, инволюирующая мезодерма перемещается по этим фибронектиновым волокнам. Это предположение было подтверждено фактом, что химически синтезированный «поддельный» фибронектин может конкурировать с фибронектином внекле-