

Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
170________________ ГЛАВА 5_______________________________________________________________________________
Таблица 5. 1. Основные производные нервного гребня 1
|
Пигментные клетки |
Сенсорная нервная система |
Вегетативная нервная система |
Скелетная и соединительная ткань |
Эндокринные железы |
|
| ||||

Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
|
| |
|
|
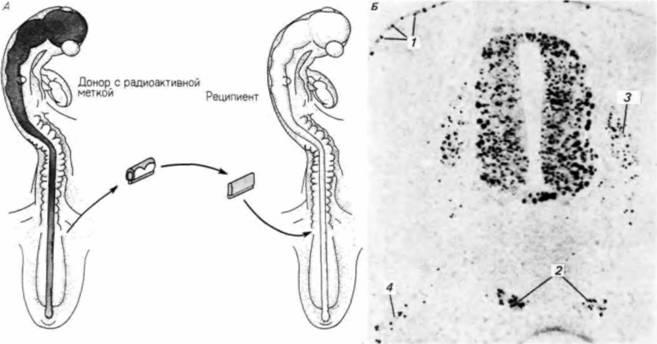
Рис. 5.31. Миграция клеток нервного гребня. А. Методика пересадки с целью маркировки клеток нервного гребня. У зародыша-донора вырезают участок дорсальных осевых структур из этого участка изолируют нервную трубку и нервный гребень и пересаживают эти структуры зародышу-реципиенту на место, из которого такие же структуры были предварительно удалены. Если клетки нервного гребня донора помечены H-тимидином или генетически (взяты от зародыша другого вида или линии), то потомки этих клеток могут быть прослежены на последующих стадиях развития. Б. Локализация методом радиоавтографии клеток нервного гребня, которые мигрировали из пересаженного донорского нервного гребня, меченного радиоактивным изотопом, и дифференцировались в меланобласты (1), клетки симпатических ганглиев (2), ганглиев дорсальных корешков (3) и глиальные клетки (4). В. Цыпленок, развившийся из зародыша непигментированной линии кур, которым был пересажен участок нервного гребня, взятый, от зародыша пигментированной линии. Из клеток пересаженного нервного гребня дифференцировались пигментные клетки, способные мигрировать в кожу крыла. (По Weston, 1963; фотографии любезно представлены J. Weston.) |

|
|
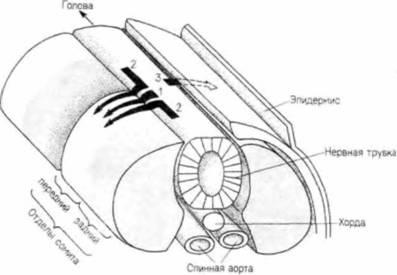
Рис. 5.32. Миграция клеток нервного гребня в туловищной области куриного зародыша. Путь 1: клетки перемещаются в вентральном направлении через передний отдел сомита. Эти клетки, вероятно, участвуют в образовании симпатических и парасимпатических ганглиев, а также становятся клетками мозгового вещества надпочечников и ганглиев дорсальных корешков. Путь 2: клетки, примыкающие к заднему отделу сомита, мигрируют между нервной трубкой и сомитом, а затем вступают в передний отдел лежащего рядом сомита. Эти клетки были обнаружены в ганглиях дорсальных корешков. Путь 3: клетки мигрируют в дорсолатеральном направлении под покровным эпителием. Эти клетки становятся меланоцитами, вырабатывающими пигмент. |
Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
172_______________ ГЛАВА 5_______________________________________________________________________________
|
|
|
|
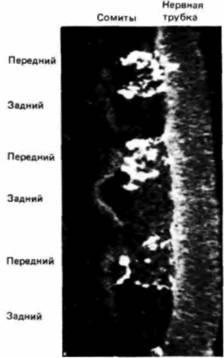
Рис. 5.33. Миграция клеток нервного гребня. Продольные срезы через сомиты, окрашенные связанной с флуоресцеином антисывороткой HNK-1, которая избирательно «узнает» клетки нервного гребня. Интенсивное свечение клеток видно только в переднем, но не в заднем отделе каждого сомита. (Из Bronner-Fraser, 1986a; фотография с любезного разрешения M. Bronner-Fraser.) |
Рис 5.34 Миграция пигментных клеток. Вид с дорсальной (А) и вентральной (Б) сторон мыши, гетерозиготной по мутации White. Меланобласты не мигрируют по всей поверхности тела мыши: в центре брюха сохраняется белое пятно (отсутствуют пигментные клетки). У мышей, гомозиготных по этой мутации (на рисунке они не показаны), шерсть белая, но в сетчатке можно видеть пигмент меланин. (Фотографии с любезного разрешения Джексоновской лаборатории.) |

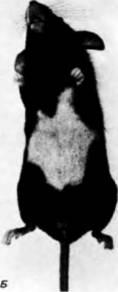
гребня, на брюхе образуется белое пятно, свидетельствующее о том, что пигментные клетки этого места не достигли (рис. 5.34).
Таким образом, клетки нервного гребня мигрируют не случайно, а следуя строго определенным путям. Природа этих направляющих их движение путей еще неизвестна, однако полагают, что миграция клеток контролируется субстратом, по которому они продвигаются. В 1941 г. Пауль Вейс высказал предположение, что между специфическими молекулами поверхности клетки и комплементарными молекулами субстрата должны возникать временные связи. Сравнительно недавно были получены данные, свидетельствующие о том, что по крайней мере в отношении клеток нервного гребня дело обстоит именно так. Во-первых, когда нервную трубку и связанный с нею нервный гребень переворачивают так, что нервная трубка оказывается вверху, клетки нервного гребня продолжают мигрировать из него. Однако теперь они движутся в дорсальном, а не в вентральном направлении, тем самым сохраняя прежнюю ориентацию по отношению к нервной трубке. Во-вторых, когда клетки нервного гребня или их производные помещают (посредством пересадки или инъекции) на место, по которому в норме пролегает путь их миграции, они мигрируют по этому пути. Другие же эмбриональные клетки неспособны ориентироваться подобным образом и остаются в том месте, на которое их поместили (Erickson et al., 1980; Bronner-Fraser, Cohen, 1980). Следовательно, клетки нервного гребня могут узнавать некоторые пути в зародыше и мигрировать по ним.
Возможно, клетки нервного гребня узнают правильный путь потому, что он маркирован белком фибронектином. Розавио и др. (Rosavio et al., 1983) было обнаружено, что мигрирующие клетки нервного гребня, прикрепляясь к этому белку, движутся по нему. Если клетки нервного гребня, помещенные в чашку Петри, приходят в контакт с полоской фибронектина, то они предпочитают двигаться по ней в направлении от более высоких концентраций клеток к более низким (рис. 5.35). Это имитирует поведение клеток нервного гребня в головной области, где такие клетки путешествуют по тонким, богатым фибронектином трекам, удаляясь от участков с высокой плотностью клеток (Mayer et al., 1981). Клетки нервного гребня обнаруживают также сродство к ламинину, другому белку, секретируемому клетками и участвующему в образовании внеклеточ-