

Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
_______________ ДРОБЛЕНИЕ: СОЗДАНИЕ МНОГОКЛЕТОЧНОСТИ 77
и перпендикулярны друг другу. Третье деление бывает экваториальным: митотические веретена в каждом бластомере располагаются параллельно анимально-вегетативной оси, и возникающие борозды отделяют полюсы один от другого, причем зародыш разделяется на восемь бластомеров равной величины. Каждый бластомер анимальной половины зародыша теперь располагается непосредственно над бластомером вегетативной половины.
Четвертое деление бывает снова меридиональным и приводит к образованию двух ярусов по 8 клеток в каждом: пятое экваториальное деление дает начало четырем ярусам, в каждом из которых содержится по 8 клеток. Далее наблюдается чередование меридиональных и экваториальных делений, приводящих к образованию 64, 128 и 256 клеток. В результате возникает зародыш, у которого бластомеры располагаются горизонтальными рядами вдоль центральной полости. На обоих полюсах зародыша бластомеры сближаются, и возникает полый шар, стенка которого образована одним слоем клеток. Такой полый шар называют бластулой, а его центральную полость - бластоцелем.
Если провести разрез через зародыш Synapta в плоскости любого меридиана на любой стадии дробления, то этот разрез разделит его на две зеркальные половины. Такой тип симметрии присущ шару или цилиндру и носит название радиальной симметрии. Соответственно мы говорим, что у Synapta дробление радиальное голобластическое.
Морские ежи
Для морских ежей также характерно радиальное голобластическое дробление, но с некоторыми существенными модификациями. Первое и второе деления дробления очень сходны с такими же делениями у Synapta; оба они меридиональные и перпендикулярны друг другу. Третье деление также экваториальное и отделяет полюсы один от другого (рис. 3.3).
|
|
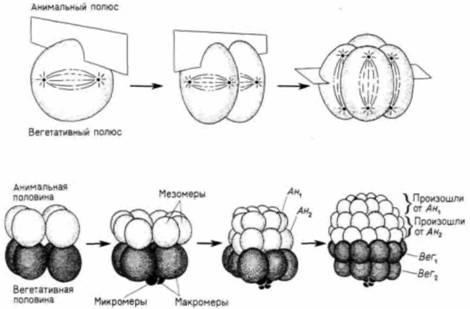
Рис. 3.3. Дробление у морского ежа. А. Плоскости деления при первых трех делениях дробления и образование отдельных ярусов клеток при 3-6 делениях. Б - Г. Микрофотографии живых зародышей морского ежа Lytechinus pictus: вид со стороны анимального полюса, Б - 2-клеточная и В - 4-клеточная стадии развития морского ежа. Г. 32-клеточная стадия представлена без оболочки оплодотворения, что позволяет различить мезомеры анимального полюса, центральные макромеры и вегетативные микромеры, которые видны в центре. (Фотографии с любезного разрешения G. Watchmaker.) |
|
| |

Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
78 ГЛАВА 3
|
|
Рис. 3.4. Образование микромеров при четвертом делении у зародышей морского ежа. Вид на вегетативный полюс зародышей снизу. А. Положение и ориентация митотического веретена в нижней части вегегативных клеток при рассматривании живого зародыша в поляризованном свете. Б. Деление в соответствии с асимметричным расположением веретен привело к образованию микромеров и микромеров. (Из Inouê. 1982: фотографии с любезного разрешения S. Inoué.) |

Четвертое деление, однако, происходит иначе, чем у Synapta. Четыре клетки анимального яруса разделяются меридионально на восемь бластомеров одинаковой величины. Эти клетки называют мезомерами. Вегетативный ярус претерпевает неравное экваториальное деление, в результате которого образуются четыре крупные клетки макромеры и четыре клетки меньшего размера микромеры у вегетативного полюса (рис. 3.4). При следующем делении восемь мезомеров 16-клеточного зародыша делятся экваториально и образуют два «анимальных» яруса, ан1, и ан2 , расположенные один над другим. Макромеры делятся меридионально, образуя восьмиклеточный ярус ниже ан2. Микромеры также делятся: возникшая маленькая группа клеток примыкает к более крупным клеткам расположенного выше яруса. При шестом делении все борозды ложатся экваториально, при седьмом меридионально. В результате седьмого деления возникает 128-клеточная бластула.
Свен Герстадиус (Hörstadius, 1939) проделал простой опыт, который показал, что время появления борозд дробления у морского ежа и их ориентация не зависят от предыдущих делений. Когда он подавлял первые одно, два или три деления, встряхивая яйца или помещая их в гипотоническую морскую воду, то неравное (четвертое) деление дробления, приводящее к образованию микромеров, все равно наступало в положенное время (рис. 3.5). На этом основании Герстадиус пришел к заключению, что существуют три фактора, которые детерминируют деление 8-клеточного зародыша: 1) «прогрессивные изменения цитоплазмы, которые определяют ориентацию веретен, образующихся спустя определенные интервалы времени после оплодотворения»: 2) в цитоплазме вегетативной области должен существовать особый материал, образующий микромеры; и 3) должен существовать какой-то механизм, который в соответствующий момент активирует материал, образующий микромеры (Hörstadius, 1973). Природа «микромерных часов» до сих пор не выяснена. Однако Икеда (Ikeda. 1965) получил данные, согласно которым клетка может вести отсчет событий, происходящих при дроблении, благодаря циклическим изменениям в цитоплазме. Он обнаружил, что освобождение сульфгидрильных (-SH ) групп белков происходит циклически. Чередование окисленной (—S—S—) и восстановленной (HS—SH) конформаций, вероятно, отражает глубокие изменения в клеточном метаболизме. Икеда показал, что свободные сульфгидрильные группы в наибольшем числе имеются в делящихся бластомерах и в наименьшем — в то время, когда клетки не делятся (рис. 3.6. А). Воздействия, которые предотвращают деления дробления, не оказывают влияния на этот цикл восстановления окисления. Отделение микромеров всегда происходит на четвертом —SHпике после оплодотворения независимо от того, какое число делений дробления было предварительно подавлено (рис. 3.6. Б). Напротив, если подавить возникновение сульфгидрильного пика (путем добавления к морской воде эфира), то отделение микромеров запаздывает. Возможно, что циклические процессы восстановления и окисления играют какую-то роль во внутриклеточном отсчете времени. Из яиц морского ежа был выделен белок (Sakai. 1966. 1968), который сокращается при окислении его свободных сульфгидрильных групп. Подобные белки, возможно, и обусловливают такие циклические изменения.
Стадия бластулы при развитии морского ежа начинается тогда, когда число клеток зародыша достигнет 128. Эти клетки образуют стенку полого шара, которая окружает центрально расположенный бластоцель (рис. 3.7). Каждая клетка находится в контакте с богатой белками жидкостью бластоцеля и с гиалиновым слоем. В это время контакт между клетками становится более тесным. Анализ этого процесса у морской звезды показал (DanSohkawa. Fujisawa. 1980), что образование полого