

Механизмы дробления Инициация дробления
Клеточный цикл соматических клеток по функциональному признаку делится на четыре стадии (или фазы). За митозом (М) следует пресинтетический период (G1), после которого происходит синтез ДНК (S). Затем наступает премитотический цикл (G2), за которым следует митоз (рис. 3.39.А). Наступление этих фаз регулируется факторами роста, которые будут подробно рассмотрены в гл. 20. Клеточный цикл бластомеров на ранних стадиях дробления может быть, однако, гораздо более простым. Бластомеры у морского ежа на этих стадиях не имеют фазы G1; репликация ДНК осуществляется в телофазе предыдущего митоза (Hinegardner et al., 1964). На тех же стадиях у Xenopus laevis и дрозофилы отсутствуют две фазы, G1 и G2. (У Xenopus laevis эти фазы появляются в клеточном цикле при переходе к средней бластуле после 12-го деления, а у дрозофилы — после 14-го. В обоих случаях это изменение цикла сочетается с увеличением клеточной подвижности и транскрипции РНК.) В течение первых 12 делений клетки Xenopus делятся синхронно и клеточный цикл бывает двухфазным: S-фаза сменяется М-фазой и M фаза —S-фазой (рис. 3.39, Б. Laskey et al., 1977; Newport, Kirschner, 1982a).
Факторы, регулирующие этот двухфазный цикл, локализованы в цитоплазме. В норме преобразования растущих ооцитов Xenopus laevis останавливаются на стадии профазы первого деления мейоза. Такие ооциты неспособны к делению. Когда в них пересаживают ядра делящихся клеток, эти ядра перестают делиться. Нормальные ооциты, подвергнутые стимулирующему воздействию прогестерона, возоб-
|
|
|
Рис. 3.39. Клеточные циклы соматических клеток и бластомеров на ранних стадиях дробления. А. Клеточный цикл типичной соматической клетки. За митозом (М) следует «интерфазное» состояние. Период между митозами подразделяется на фазы G1, S (синтез) и G2. Б. Более простой двухфазный цикл бластомеров амфибий на ранних стадиях дробления, имеющий только две фазы S и M. |

Гилберт с.Биология развития: в 3-х т. Т. I: Пер. С англ. — м.: Мир, 1993. — 228 с.
102 Глава 3
новляют мейотические преобразования, которые идут до стадии метафазы второго деления мейоза и на этой стадии останавливаются. Если в цитоплазму обработанных прогестероном ооцитов помещают ядра неделящихся клеток (например, нейронов), то такие ядра тоже инициируют деление (Gurdon. 1968). В цитоплазме происходят периодические сокращения кортикального слоя (характерные для деления) даже при отсутствии ядер или центриолей. Если в энуклеированные зародыши инъецируют клонированные фрагменты ДНК, то их репликация происходит под контролем этих циклов (Нага et al.. 1980; Harland. Laskey. 1980: Karsenti et al.. 1984). Таким образом, способность клетки к делению регулируется цитоплазмой.
Недавно были выявлены факторы, осуществляющие эту регуляцию. Фактор, индуцируемый прогестероном, который позволяет ооциту возобновить мейоз, представляет собой белок с молекулярной массой 100 000 дальтон: его называют фактором, стимулирующим созревание (maturation promoting factor, MPF). После первого деления мейоза повышается концентрация другого белка цитостатического фактора (cytostatic factor, CSF). что приводит к остановке преобразований ооцита на метафазе второго деления мейоза. Ионы кальция, высвобождающиеся при оплодотворении из эндоплазматического ретикулума. по-видимому, инактивируют CSF, что делает возможным завершение мейоза и слияние пронуклеусов (Meyerhof, Masui. 1977).
Те же самые факторы— MPF, CSF и ионы кальция — важны и после оплодотворения, поскольку они регулируют двухфазный клеточный цикл на ранних стадиях дробления Xenopus. Показано (Gerhart et al.. 1984), что в делящихся клетках уровень активности MPF претерпевает циклические изменения. Активность MPF в бластомерах лягушки на ранних стадиях дробления наивысшая в М-фазе и не выявляется в S-фазе. Такая же цикличность обнаруживается и в энуклеированных бластомерах. Установлено (Newport. Kirschner, 1984), что даже при отсутствии белкового синтеза наступление репликации ДНК (Sфаза) и вступление в митоз (М-фаза) определяются исключительно увеличением и утратой активности MPF. Дробящиеся клетки могут быть блокированы в S-фазе путем инкубации их с ингибитором белкового синтеза. Когда в такие клетки инъецируют MPF, они вступают в М-фазу. Ядерная оболочка этих клеток разрушается, и происходит конденсация хроматина с образованием видимых хромосом. Через 1 ч активность MPF снижается и хромосомы возвращаются к состоянию, характерному для S-фазы. MPF действует, вероятно, путем изменения структуры ядерной оболочки. Через 15 мин после его введения наблюдается гиперфосфорилирование трех ос
|
|
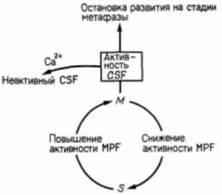
Рис. 3.40. Модель регуляции клеточного цикла бластомеров на ранних стадиях дробления у амфибий В присутствии MPF S-фаза клеточного цикла переходит в М-фазу. Когда активность ΜPF прекращается (в результате его разрушения или функциональной инактивации), совершается переход от М-фазы к S-фазе. CSF стабилизирует MPF, задерживая клетку в М-фазе. Ионы кальция противодействуют эффекту CSF, т.е. они возвращают клетку в фазу S. (По Newport, Kirschner, 1984.) |
новных белков ядерной оболочки. В течение следующих 15 мин оболочка деполимеризуется и разрушается (Miake-Lye, Kirschner, 1985).
Модель регуляции двухфазного клеточного цикла представлена на рис. 3.40. В цитоплазме продолжается образование MPF (либо путем трансляции мРНК для MPF, уже запасенной в клетке, либо путем активации предшественника MPF). MPF побуждает клетку к переходу от S-фазы к M-фазе. Когда MPF инактивируется, клетка возвращается в S-фазу. Таким образом. MPF управляет клеточным циклом. CSF и ионы кальция регулируют работу регулятора, контролируя содержание MPF в клетке. CSF, по-видимому, стабилизирует MPF, удерживая клетку в состоянии митоза. Ионы кальция, инактивируя CSF, тем самым создают возможность инактивации MPF. При добавлении CSF прекращаются также циклические сокращения кортикального слоя цитоплазмы, а последующая инъекция ионов кальция снова их стимулирует. Таким образом, на ранних стадиях развития цитоплазма определяет скорость клеточных делений и продолжительность S- и М-фаз цикла.