

Контрольные вопросы и задания
1. Какие причины вызывают речевые расстройства?
2. Рассмотрите клинико-педагогические классификации нарушений речи и свяжите их с локализацией патологического процесса.
3. Какие основные клинические формы нарушений речи периферического характера требуют участия логопеда?
4. Перечислите нарушения речи центрального происхождения и составьте таблицу нарушений.
5. На каких принципах основана психолого-педагогическая классификация нарушений речи? Какие существуют способы помощи детям с нарушениями речи?
Раздел IV зрительная система
Глава 15 строение зрительной системы
15.1. Краткие сведения об эволюции зрительной системы
Зрение — один из важнейших органов чувств человека. Оно эволюционно приспособлено к восприятию узкой части диапазона электромагнитных излучений (видимый свет). Зрительная система дает мозгу около 80 % сенсорной информации.
Зрение — это многозвеньевой процесс, который начинается с проекции изображения на сетчатку глаза, возбуждения фоторецепторов, передачи и преобразования зрительной информации в нейронных слоях зрительной системы, а заканчивается принятием высшими корковыми отделами зрительной системы решения о зрительном образе.
По данным ВОЗ, в настоящее время в мире насчитывается 150 млн. лиц со значительными зрительными расстройствами, в том числе около 42 млн. слепых. За последние 15 лет численность незрячих увеличилась на 12 млн. человек, каждый четвертый из них — ребенок или человек, утративший зрение в детстве. Распространенность инвалидности вследствие патологии органа зрения у населения РФ довольно высока: она составляет 28,8 человек на 10000 взрослого населения. Уровень детской инвалидности по зрению — 5,2 человека на 10 000 населения соответствующего возраста.
Таким образом, число инвалидов по зрению в России за последнее десятилетие значительно возросло. Зрительные расстройства у каждого второго инвалида по зрению в возрасте 19 —50 лет возникли в детском возрасте. Поэтому изучение строения и функций зрительной системы, а также различных заболеваний органа зрения, приводящих к стойким нарушениям зрительного восприятия, является важным направлением в подготовке специальных педагогов и психологов.
Эволюция зрительных приспособительных механизмов и зрительных ощущений у различных живых существ идет разными путями. Так, например, амеба реагирует на свет изменением движения и укорочением обращенных к свету псевдоподий, что, по-видимому,
обусловлено изменением химического состава и физического состояния протоплазмы. У гидры, так же как и у амебы, нет специфических светочувствительных клеток, однако в результате изменений в протоплазме под влиянием света гидра изменяет движение.
Первые более или менее структурно оформленные аппараты для восприятия света встречаются у простейших. Это скопления мелкозернистого пигмента, образующего стигму, или глазное пятно. У эвглены, например, такое скопление лежит у основания жгутика и тесно связано с базальными тельцами, которые являются «генераторами» движения последнего (рис, 77). Поскольку тело эвглены не совсем прозрачно, стигма освещена сильнее, когда жгутик направлен к свету. В связи с этим жгутиконосцы проявляют положительный фототропизм.
У многоклеточных животных световоспринимающие аппараты известны начиная с кишечнополостных. Например, реакция на свет у дождевого червя - это уже не автоматическая реакция протоплазмы.
У червя имеются рассеянные по всему телу примитивные световоспринимающие (зрительные) клетки. Некоторое структурное усовершенствование наблюдается у пиявки. На ее головной части имеется группа светочувствительных клеток, окруженных пигментом, который, как известно, хорошо поглощает свет, и, следовательно, усиливает раздражение зрительных клеток.
Следующая ступень в развитии зрительных приспособлений — это глазные ямки со светочувствительными клетками, окруженными пигментом, у морских звезд и улиток.
На ранних стадиях эволюции многоклеточных развитие зрительной системы пошло по двум основным путям. Первый путь был связан с возрастанием числа световоспринимающих элементов. Это увеличило коэффициент поглощения светового потока и расширило сферу зрительного восприятия.
Наиболее сложными устройствами данного типа являются фасеточные глаза ракообразных и насекомых. Они состоят из многих (от сотен до десятков тысяч) отдельных светочувствительных элементов, называемых омматидиями. Типичный омматидий — это узкий усеченный конус, содержащий совокупность светопреломляющих, светочувствительных и



светоизолирующих элементов (рис. 78). Если свет падает на омматидий вдоль его оси, то светопреломляющий аппарат омматидия, состоящий из выпуклой линзы (хрусталика) и кристаллического конуса, концентрирует световой поток на светочувствительной клетке омматидия — рабдоме.
Если же падающий луч составляет с осью омматидия угол больше 2—5°, то большая часть его не попадает на рабдом, а поглощается темным пигментом, изолирующим омматидий от соседних. Благодаря этому свойству глаза некоторых насекомых имеют достаточно высокую разрешающую способность: острота зрения может достигать 0,1—0,2°. Поле зрения обоих сложных глаз у многих видов членистоногих охватывает почти полную сферу.
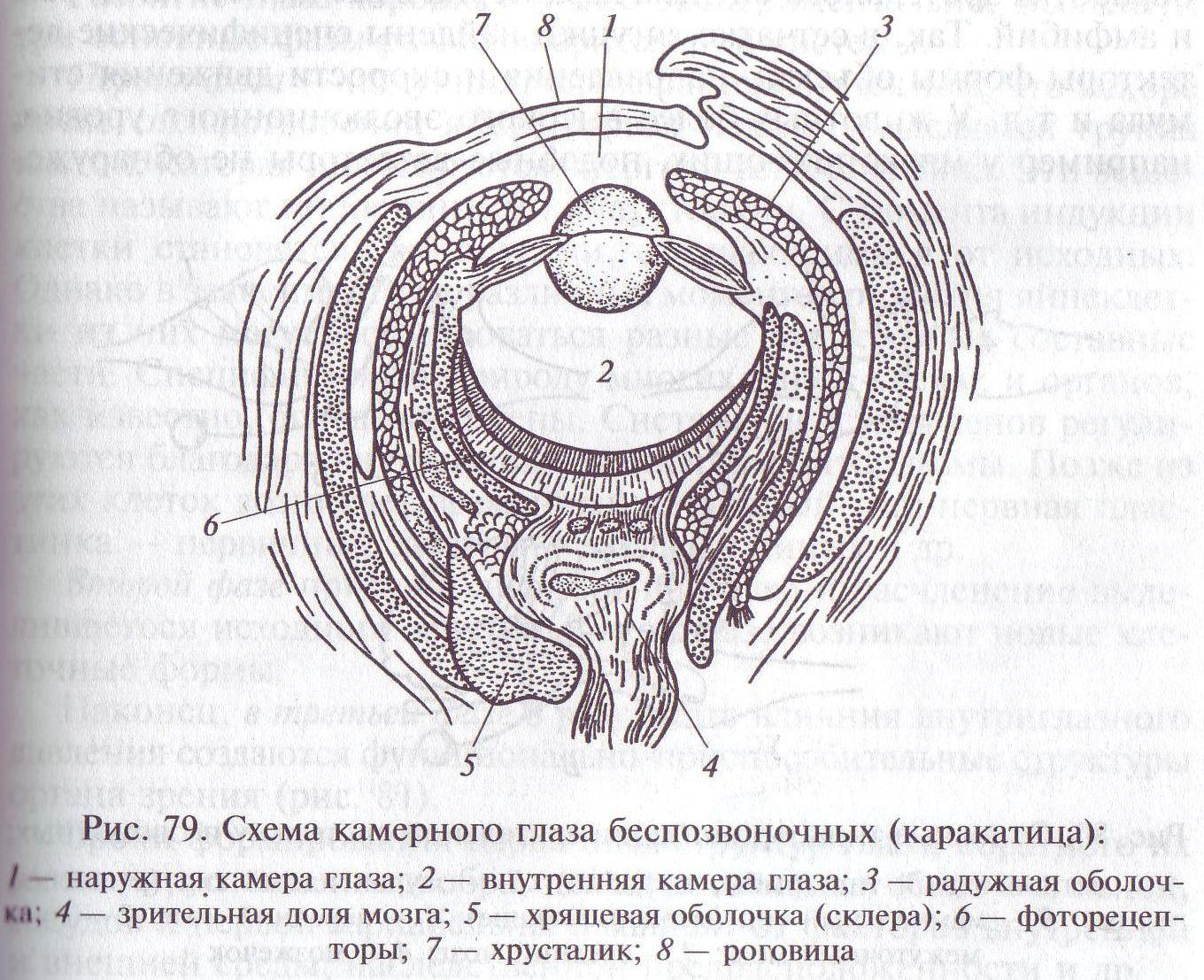
Второй путь развития светочувствительного аппарата состоял в формировании единой для всего глаза оптической системы (рис. 79). Наиболее примитивным органом зрения подобного рода являются светочувствительные органы некоторых кишечнополостных, а также личинок ряда насекомых. Они расположены на дне более или менее глубокой ямки и защищены от бокового света непрозрачными стенками (см. рис. 79), что позволяет определять направление лучей света от каждого из достаточно удаленных друг от друга источников.
В процессе филогенетического развития концевые световоспринимающие аппараты светочувствительных элементов оказывались повернутыми к свету (конвертированное устройство зрительного аппарата).
Более совершенное устройство — обскура с маленьким отверстием и внутренней светочувствительной полостью имеется у моллюска. Световоспринимающие элементы оказываются повернутыми внутрь, т.е. от света (инвертированный тип глаза) (рис. 79). Разрешающая сила, т. е. способность воспринимать близкие друг к другу мелкие детали порознь, у такого глаза много выше, чем у
глазной ямки с широким отверстием. В то же время при узком отверстии изображение на сетчатке содержит больше деталей, хотя она воспринимает гораздо меньший поток света, чем при широком отверстии.
Уже на ранних ступенях эволюции намечается разрешение этого противоречия. Так, у некоторых медуз и червей полость глазной ямки заполняется прозрачным выделением эктодермы. Часть содержимого принимает вид двояковыпуклой линзы — хрусталика, имеющего больший показатель преломления по сравнению с окружающей средой. Свет, попадающий в такой глаз от небольших объектов, концентрируется хрусталиком на небольшую часть светочувствительной поверхности сетчатки. В глазах такой конструкции освещенность изображения источника света на сетчатке может быть во много раз больше освещенности, создаваемой тем же источником на наружной поверхности глаза.
Глаза у позвоночных животных устроены по второму типу, т. е. имеют развитый оптический аппарат. Однако в отличие от всех других типов животных их сетчатка имеет гораздо более сложное строение и ряд особенностей. Во-первых, для позвоночных характерно своеобразное расположение светочувствительных клеток, которые обращены не к световому потоку, а в противоположную сторону. Таким образом, лучи света возбуждают фоторецепторы
не непосредственно, а лишь проходя через оптически прозрачные среды сетчатки и отражаясь клетками пигментного эпителия. Такое явление не наблюдается у других групп животных, и физиологическая роль его до сих пор не вполне понятна.
Во-вторых, кроме клеток, выполняющих непосредственно световоспринимающую функцию (палочки и колбочки), у позвоночных имеются нейроны 2-го порядка (так называемые биполярные клетки), которые связаны через синапсы с одним или несколькими фоторецепторами. Нейроны 3-го порядка (ганглиозные клетки) интегрируют сигналы от одного или нескольких биполяров и являются «выходными» элементами сетчатки, которые формируют сигнал, адресованный высшим отделам зрительной системы.
На уровне между фоторецепторами, биполярами и ганглиозными клетками сетчатки находятся элементы с горизонтальным расположением Волокон. Связывая упорядоченным образом нейроны сетчатки друг с другом, они формируют специфическую форму зрительных рецептивных полей. Таким образом, в сетчатке позвоночных происходит весьма сложная обработка зрительного сигнала, которая выражается в детектировании (обнаружении) тех или иных свойств действующего раздражителя. Сложная обработка зрительного сигнала характерна даже для сетчатки рыб и амфибий. Так, в сетчатке лягушки найдены специфические детекторы формы объекта, направления и скорости движения стимула и т.д. У животных более высокого эволюционного уровня, например у млекопитающих, подобные детекторы не обнаружены

т.е. в функциональном отношении их сетчатка устроена прошепчем у амфибий. Эти функции принимают на себя центральные структуры зрительного анализатора, в частности первичная зрительная кора.
По строению центральных звеньев зрительной системы всех позвоночных можно подразделить на три группы: животные с ихтиопсидным, зауропсидным и маммальным типом организации зрительного анализатора (рис. 80). Для ихтиопсидпого типа, к которому относятся рыбы и амфибии, характерно то, что окончательная обработка зрительной информации осуществляется на уровне среднего мозга (крыша мезенцефалона и претектальные ядра) и лишь незначительная часть информации поступает в передний мозг. Для зауропсидного типа (рептилии и птицы) характерно формирование нового анатомического пути через промежуточный мозг (таламус) к коре. Для маммального типа (млекопитающие) этот путь становится основным, и лишь небольшая часть зрительной информации перерабатывается на мезенцефальном уровне.