

3.3. Центральный отдел слуховой системы
3.3.1. Подкорковые центры слуха
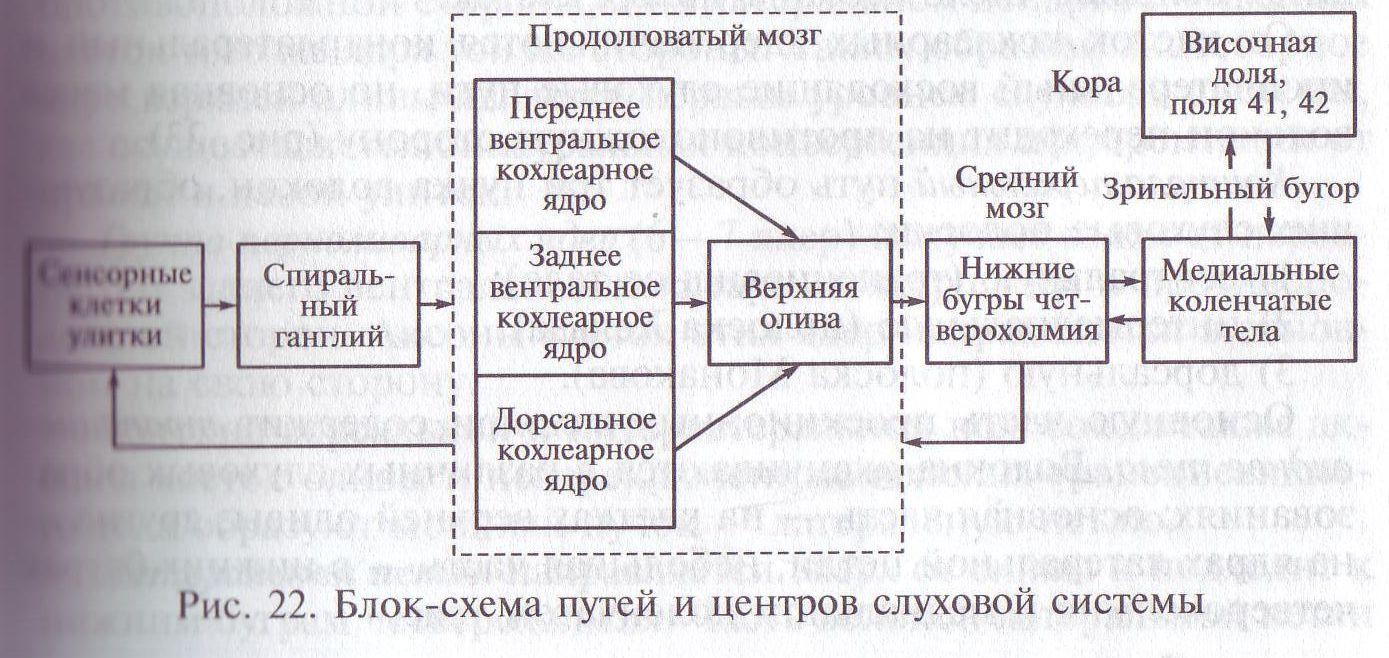
В работах С. Рамон-и-Кахаля дано описание классического слухового пути. Этот восходящий специфический (или афферентный) путь представляет собой несколько последовательных уровни (рис. 22):
1. спиральный ганглий улитки;
2. кохлеарные ядра продолговатого мозга;
3. верхняя олива продолговатого мозга;
4. нижние бугры четверохолмия среднего мозга;
5. медиальные коленчатые тела таламуса;
6. слуховые поля височной коры головного мозга.
Кроме классического пути были обнаружены дополнительные водящие слуховые пути, связанные с ретикулярной формацией ствола, мозжечком, ассоциативными ядрами таламуса и корковыми полями, окружающими центральное слуховое поле. Наряду с восходящими путями существует также система нисходящих слуховых путей, идущих от слуховой коры к нижележащим слуховым
образованиям. Слуховая система имеет многочисленные связи с другими сенсорными системами и моторными центрами.
Спиральный ганглий улитки и кохлеарный нерв. Первый уровень в восходящем слуховом пути составляют клетки спирального ганглия улитки, являющиеся нейронами I порядка. Эти биполярные (с двумя отростками) клетки, расположенные в виде тяжа плотно упакованных компактных групп, находятся в спиральном канале улитки параллельно органу Корти. Спиральный ганглий по длине значительно меньше, чем орган Корти, и делает меньше оборотов вокруг оси улитки. Тела всех клеток спирального ганглия и их отростки покрыты миелиновыми оболочками (как телефонный кабель).
Периферические отростки клеток спирального ганглия направляются к органу Корти, а центральные выходят во внутренний слуховой проход и образуют кохлеарный (или слуховой) нерв.
Кохлеарные ядра продолговатого мозга. В них переключаются все волокна слухового нерва. Он представляет собой слуховую ветвь VIII пары черепно-мозговых нервов. Слуховой нерв входит в продолговатый мозг и оканчивается на клетках кохлеарных ядер. В кохлеарном комплексе выделяют три основные ядра различной структуры: 1. переднее вентральное; 2. заднее вентральное; 3. дорсальное (или слуховой бугорок).
Ядра представляют собой скопление нейронов II порядка. Волокна слухового нерва при входе в кохлеарные ядра разделяются на две ветви — нисходящую и восходящую, иннервирующие соответственно клетки заднего и переднего ядер. Волокна от апикальной части улитки иннервируют вентролатеральную часть кохлеарного комплекса, а от базальной части — его дорсомедиальную область.
Дорсальное ядро кохлеарного комплекса по структуре резко отличается от вентральных ядер слоистым строением, сходным с корой, и сложным характером внутренних связей, а также обилием разветвлений нисходящих волокон. В кохлеарных ядрах находится большое число ассоциативных клеток.
От клеток кохлеарных ядер начинаются контрлатеральный и ипсилатеральный восходящие слуховые пути, но основная масса волокон переходит на противоположную сторону (рис. 23).
Контрлатеральный путь образует три пучка волокон, образующих слуховые полоски:
1. вентральную (трапециевидное тело); 2. интермедиальную (полоска Хельда); 3. дорсальную (полоска Монакова).
Основную часть проекционных волокон содержит трапециевидное тело. Волокна оканчиваются в различных слуховых образованиях: основная часть — на клетках верхней оливы, другие — на ядрах латеральной петли, небольшая часть - в нижних буграх четверохолмия и медиальном коленчатом теле.

Верхняя олива. В комплекс ядер верхней оливы входят: 1) S-образное (латеральное) ядро; 2) медиальное (акцессорное) ядро; 3) группа периоливарных ядер.
S-образное ядро получает афферентацию только от ипсилатерального кохлеарного ядра, в то время как аксоны его клеток иннервируют выше лежащие ядра как своей, так и противоположной стороны.
Клетки акцессорного ядра имеют два дендрита; на медиальном дендрите оканчиваются аксоны клеток переднего кохлиарного ядра противоположной стороны, а на латеральном дендрите — аксоны клеток того же ядра той же стороны. Таким образом, акцессорное ядро верхней оливы является первым уровнем слуховой системы, осуществляется бинауральная конвергенция афферентации от правой и левой улиток.
Группа периоливарных ядер (6 — 7 ядер) получает связи от переднего заднего вентральных кохлеарных ядер своей и противоположной сторон. Аксоны клеток этих ядер направляются в основан на свою сторону.
Волокна дорсальной и интерлатеральной полосок, а также аксоны клеток оливы и непереключенные волокна трапециевидного тела образуют мощный пучок — латеральную петлю.
Латеральная петля направляется вверх от оливы и подходит к нижним буграм четверохолмия. Часть волокон латеральной петли
оканчивается на клетках заднего бугра, а другая — в таламусе. Небольшое число волокон латеральной петли направляется на противоположную сторону, образуя комиссуру Пробста. Латеральная петля представляет собой общий путь для аксонов нейронов II, III и отчасти IV порядков и представляет улитку как своей, так и противоположной стороны. Латеральная петля — основной коллектор аксонов слуховых ядер продолговатого мозга.
Нижние бугры четверохолмия — среднемозговой центр слуховой системы (иначе называется задним, или нижним, двухолмием) состоит из нейронов III, IV, V порядков. Структура этого образования очень сложная. Основная часть клеточных элементов заднего холма расположена в центре и составляет его центральное ядро. Аксоны клеток заднего холма направляются к медиальному коленчатому телу своей стороны и частично к противоположному заднему холму.
Медиальные коленчатые тела — таламический центр слуховой системы. В нем различают крупно- и мелкоклеточную (главную) части. Мелкоклеточная часть, через которую проходит основной слуховой путь, неоднородная и подразделяется на дорсальную, вентральную и медиальную области. Для дорсальной области характерны крупные нейроны с толстыми дендритами, отходящими от тела клетки в различных направлениях и образующими сферическое дендритное поле. Для вентральной области основными являются нейроны с пучковидными дендритами, которые отходят от тела клетки только в одной плоскости, образуя дендритное поле дисковидной формы.
В медиальной области встречаются нейроны обоих типов, а также нейроны промежуточной формы.
Вентральная область имеет наибольшие размеры, и в ней оканчивается большая часть входящих волокон. Здесь была обнаружена ламинарная структура (область синаптических контактов), причем клетки каждого слоя иннервируются афферентными волокнами с двух сторон. Аксоны клеток медиального коленчатого тела образуют акустическую радиацию и оканчиваются на клетках слуховой коры. Комиссура Гуддена связывает оба коленчатые тела.
Итак, путь электрических импульсов от рецепторов органа слуха к слуховой коре больших полушарий головного мозга имеет 3 — 5 уровней переключения (переключательных станций) и не менее трех перекрестов (рис. 24).
Информация о звуковом потоке, попадающем в диапазон возможностей рецепторной части органа слуха по аксонам нервных клеток слухового спирального ганглия, дендриты которых подходят к рецепторным клеткам, передается в слуховой центр продолговатого мозга (кохлеарные ядра) в форме коротких электрических импульсов. Они распространяются вдоль волокон слухового нерва со скоростью от 0,5 до 100 м/с — в зависимости от толщины волокон.
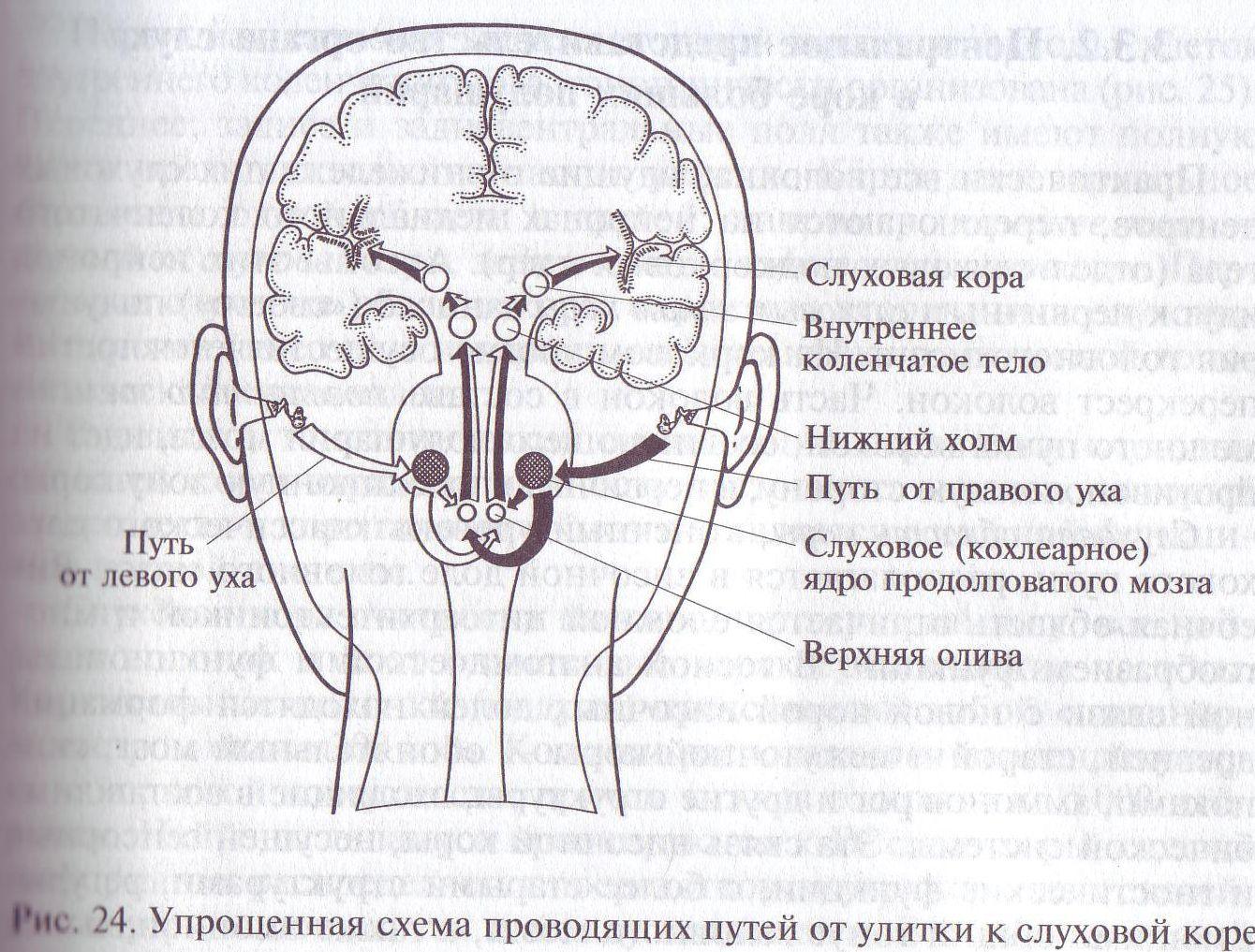
После переключения волокон слухового нерва на клетках кохлеарных ядер продолговатого мозга электрические импульсы поступают к следующему клеточному скоплению – ядрам верхней оливы. Здесь отмечается первый перекрёст слуховых путей: меньшая часть волокон остаётся в пределах полушария, на стороне которого расположены периферические слуховые рецепторы, а большая часть идет в противоположное полушария головного мозга. В области основания мозга, где располагается перекрест, имеется еще одна группа ядер — ядра трапециевидного тела. В них также осуществляется частичное переключение волокон клеток кохлеарных ядер. Небольшая часть этих волокон направляется, переключаясь, в средний мозг и заканчивается на клетках нижних (задних) холмов. Сюда же приходит значительная часть перекрещенных и не перекрещенных волокон из ядер верхней оливы.
Подавляющее большинство волокон от клеток кохлеарных ядер, верхнеоливарных ядер, трапециевидного тела и латеральной петли переключается на клетках нижних холмов, после чего волокна следующего порядка либо переходят в противоположное полушарие (второй крупный перекрест), либо идут непосредственно к ближайшим подкорковым слуховым центрам таламуса (зрительного бугра) — медиальным коленчатым телам. Только очень небольшая волокон проходит мимо нижних холмов, не переключаясь в них, и заканчивается прямо в медиальном коленчатом теле.