Клонирование генов |
|
|
|
|
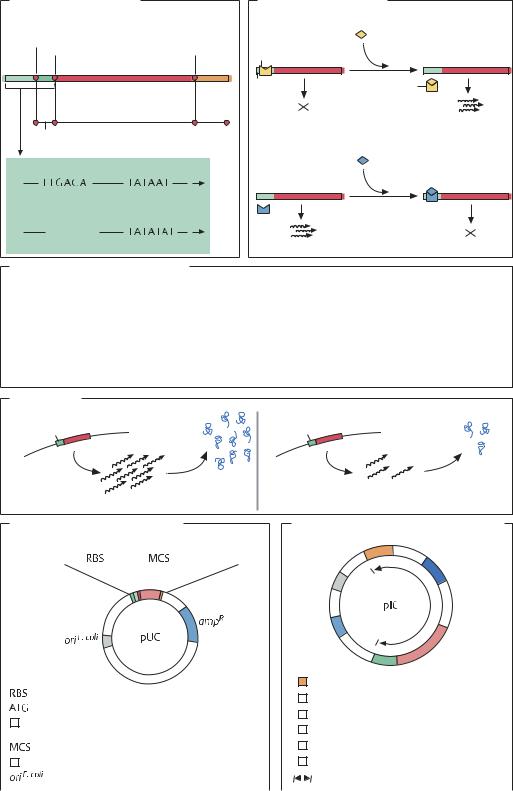
Методы идентификации |
|
|
|||
|
a |
Проба ДНК |
б |
|
Саузерн-блот |
Поиск ДНК |
|
Гибридизация |
||
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
с меченым ДНК- |
Получение библиотеки генов |
|
ПЦР |
|
|
|
|
|
или РНК-зондом |
||
|
|
|
|
|
|
|
||||
Гибридизация для поиска |
|
Секвени- |
|
Нозерн-блот |
Поиск мРНК |
|
Гибридизация |
|||
|
нужного клона |
|
|
рование |
|
|
|
|
|
с меченым ДНК- |
|
|
|
|
гена |
|
|
|
|
|
или РНК-зондом |
|
|
|
|
|
|
|
|
|
|
|
|
Очистка ДНК |
|
|
|
|
Вестерн-блот |
Поиск белка |
|
Иммуноанализ |
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
с мечеными |
|
Получение |
|
|
|
|
|
|
|
|
антителами |
|
|
|
|
|
|
|
|
|
|
|
коротких фрагментов гена |
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
Использование |
Поиск |
|
Экспрессия |
|
|
|
|
|
|
|
репортерных |
регуляторных |
|
репортерных |
|
Секвенирование гена |
|
|
|
групп |
элементов |
|
белков |
|||
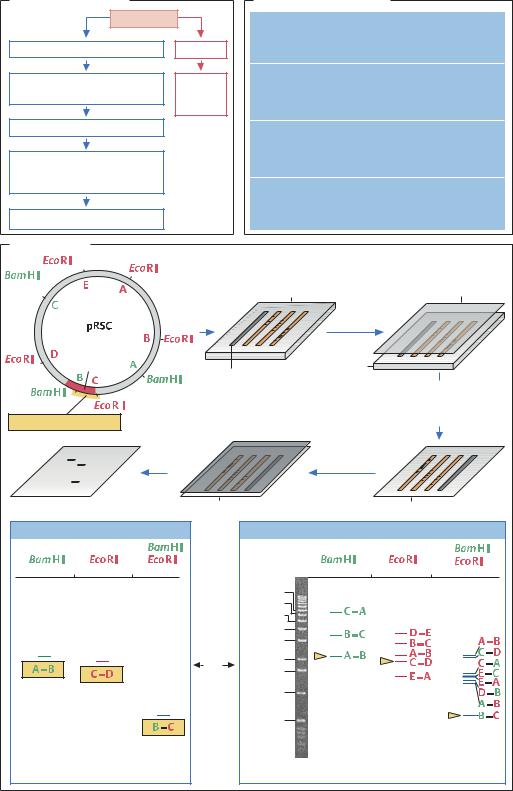
Саузерн-блот |
|
|
|
|
|
|
|
|
|
|
|
11 540 п.н. |
997 п.н. |
Расщепление рестриктазами |
|
|
|||||
|
|
|
|
Нейлоновая |
||||||
10 068 п.н. |
|
|
|
BamHI и EcoRI, электрофорез |
|
|||||
|
|
|
в агарозном геле |
|
|
мембрана |
||||
|
|
|
|
|
|
|
|
Перенос |
|
|
|
12 002 п.н. |
|
|
|
|
|
|
|
|
|
|
|
|
3 188 п.н. |
|
|
|
|
|
|
|
7 959 п.н. |
Ген |
|
|
|
|
|
Агарозный гель |
|
|
|
|
|
|
Маркерная ДНК |
|
|
|
||||
|
|
4 467 п.н. |
|
|
|
|
Гибридизация |
|||
|
|
|
|
|
со специфическим зондом |
|||||
6 621 п.н. |
6 022 п.н. |
|
|
|
|
|||||
|
|
|
|
|
(содержащим |
|||||
Специфический зонд |
|
|
|
|
|
|
радиоактивную метку) |
|||
|
|
|
|
Радиоавтография |
|
|
|
|||
|
|
|
|
|
|
|
|
|
||
|
|
|
Рентгеновская |
|
|
|
|
|
|
|
|
|
|
пленка |
|
|
|
|
|
|
|
Проявленная пленка |
|
|
Нейлоновая мембрана |
Нейлоновая мембрана |
||||||
|
Рентгеновская пленка |
|
|
|
|
|
Агарозный гель |
|
||
|
|
|
|
|
|
|
Маркерная |
|
|
|
|
|
|
|
|
|
|
ДНК |
|
|
|
|
|
|
|
|
|
|
7 |
|
|
|
|
|
|
|
|
|
|
6 |
|
|
|
|
|
|
|
|
|
|
5 |
|
|
|
|
|
|
|
|
|
|
4 |
|
|
|
|
|
|
|
|
|
|
3 |
|
|
|
|
|
|
|
Срав- |
н. |
2 |
|
|
|
|
|
|
|
|
не- |
|
т. п. |
|
|
|
|
|
|
|
|
|
1,6 |
|
|
|
||
|
|
|
|
ние |
|
|
|
|
||
|
|
|
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
500 |
|
|
|
|
Следы |
|
|
|
|
|
|
|
Следы |
|
1 |
2 |
|
3 |
|
|
|
Маркер |
1 |
2 |
3 |
245