

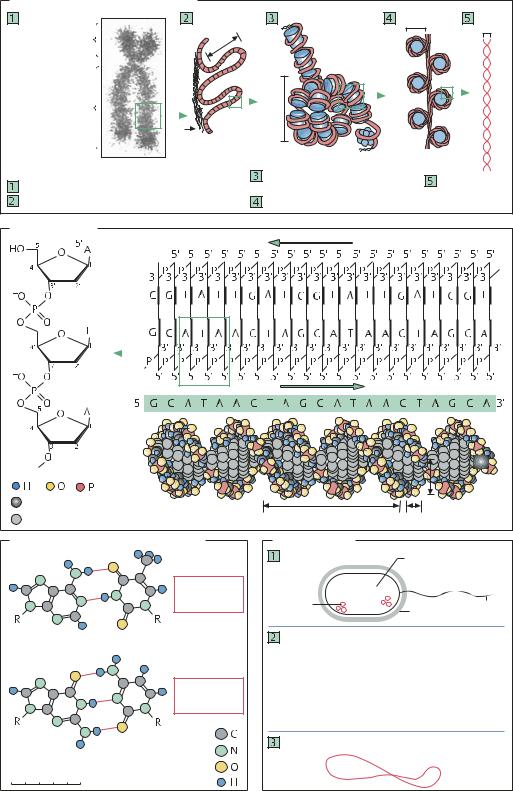
От хромосомы к двойной спирали |
|
|
|
250 нм |
10 нм |
2 нм |
|
Короткое плечо (p) |
|
|
|
Центромера |
|
|
|
Длинное плечо (q) |
нм |
|
|
30 |
|
||
|
|
||
Гистон |
|
|
|
|
Хроматиновая фибрилла, |
Двойная |
|
Целая метафазная хромосома |
содержащая 1200 п.н. |
||
спираль ДНК – |
|||
|
|
||
Петли хроматина – около 100 000 п.н. на петлю |
«Бусины» нуклеосом – по 80 п.н. |
10 п.н. в 3,4 нм |
|
Структура ДНК |
|
|
С в сахарофосфатном остове |
|
1 нм |
|
|
С или N в азотистых основаниях |
3,4 нм |
0,34 нм |
|
|
|
|
|||
Комплементарные пары оснований |
ДНК бактерий |
|
|
|
в ДНК |
|
Бактериальная клетка |
Нуклеоид, содержащий |
|
|
|
|||
|
|
|
суперскрученную |
|
|
Пара АТ – два |
Плазмид- |
молекулу ДНК |
|
|
водородных |
|
|
|
|
мостика |
ная ДНК |
Клеточная |
Жгутик |
|
|
|
стенка |
|
Аденин (А) |
Тимин (Т) |
Раскручивание витков спирали в результате |
||
|
|
разрывов, образованных ферментом ДНКазой I |
||
|
Пара GС – три |
|
|
|
|
водородных |
|
|
|
|
мостика |
|
|
|
|
|
Открытая кольцевая форма молекулы ДНК |
|
|
Гуанин (G) |
Цитозин (С) |
|
|
|
0 |
0,5 нм |
|
|
225 |
|
|
|
|
|

Функции ДНК |
|
|
|
|
ДНК |
Транскрипция |
Трансляция** |
Белок |
Рибосомы на мРНК |
Обратная |
мРНК |
|
||
|
|
|
|
|
|
транскрипция* |
|
|
|
Репли- |
|
|
|
|
кация |
|
|
|
|
|
|
|
ДНК |
РНК-полимераза |
ДНК |
Наследование |
|
0,5 мкм |
|
|
|
|||
* только у ретровирусов |
|
Транскрипция бактериальной ДНК |
||
** на рибосомах |
|
|
|
|
Генетический код |
|
|
|
|
Аминокислота |
Кодон |
Аминокислота |
Кодон |
Аминокислота |
Кодон |
Аминокислота |
Кодон |
|
Фенилаланин (F) |
UUU |
Серин (S) |
UCU |
Тирозин (Y) |
UAU |
Цистеин (C) |
UGU |
|
|
|
UUC |
|
UCC |
|
UAC |
|
UGC |
Лейцин (L) |
UUA |
|
UCA |
Стоп-кодон |
UAA |
Стоп-кодон |
UGA |
|
|
|
UUG |
|
UCG |
Стоп-кодон |
UAG |
Триптофан (W) |
UGG |
|
|
|
|
|
|
|
|
|
Лейцин (L) |
CUU |
Пролин (P) |
CCU |
Гистидин (H) |
CAU |
Аргинин (R) |
CGU |
|
|
|
CUC |
|
CCC |
|
CAC |
|
CGC |
|
|
CUA |
|
CCA |
Глутамин (Q) |
CAA |
|
CGA |
|
|
CUG |
|
CCG |
|
CAG |
|
CGG |
|
|
|
|
|
|
|
|
|
|
Изолейцин (I) |
AUU |
Треонин (T) |
ACU |
Аспарагин (N) |
AAU |
Серин (S) |
AGU |
|
|
AUC |
|
ACC |
|
AAC |
|
AGC |
|
|
AUA |
|
ACA |
Лизин (K) |
AAA |
Аргинин (R) |
AGA |
Метионин (M) |
AUG |
|
ACG |
|
AAG |
|
AGG |
|
|
|
|
|
|
|
|
|
|
|
Валин (V) |
GUU |
Аланин (A) |
GCU |
Аспарагиновая кислота (D) GAU |
Глицин (G) |
GGU |
|
|
|
GUC |
|
GCC |
|
GAC |
|
GGC |
|
|
GUA |
|
GCA |
Глутаминовая кислота (E) GAA |
|
GGA |
|
|
|
GUG |
|
GCG |
|
GAG |
|
GGG |
|
|
|
|
|
|
|
|
|
Создание праймеров |
|
Частота использования |
|
|||
|
|
кодонов некоторых |
|
|||
|
Выделенный белок |
аминокислот у различных |
||||
|
|
организмов |
|
|
|
|
|
Определение аминокислотной |
Амино- |
Кодон |
Вероятность |
||
|
последовательности белка |
|||||
|
кислота |
|
использования |
|||
|
или его участка |
|
||||
|
|
|
|
кодона, % |
||
|
Информация |
Glu, E |
GAG |
0,30 |
0,31 |
0,59 |
|
|
GAA |
0,70 |
0,69 |
0,41 |
|
|
о последовательности белка |
|
||||
|
|
Lys, K |
AAG |
0,24 |
0,43 |
0,60 |
|
|
|
AAA |
0,76 |
0,57 |
0,40 |
Синтез набора |
Набор всех возможных |
Pro, P |
CCG |
0,55 |
0,12 |
0,11 |
«вырожденных» |
последовательностей ДНК, |
|
CCA |
0,20 |
0,42 |
0,27 |
праймеров |
кодирующих изучаемый |
|
||||
Гибридизация |
полипептид |
|
CCU |
0,16 |
0,31 |
0,29 |
с банком ДНК |
N = любое основание |
|
||||
или кДНК |
|
CCC |
0,10 |
0,15 |
0,33 |
|
|
|
|
||||
|
Определение истинной |
|
|
|
|
|
|
последовательности кДНК |
E. coli |
|
|
|
|
|
|
|
|
|
|
|
|
|
S. cerevisiae |
|
|
|
|
|
|
Человек |
|
|
|
|
227