

инженерии |
Эксперимент в генетической инженерии |
||
ВВЕДЕНИЕ. В генно-инженерных исследованиях при- |
меняют обратные транскриптазы с различными свой- |
||
|
|||
|
меняются очень многие методы. К наиболее часто |
ствами. Чаще всего используют ДНК-полимеразу, |
|
генетической |
используемым относятся следующие: 1) выделение |
выделенную из Thermus aquaticus. Этот фермент, ус- |
|
ДНК, ее амплификация, различные модификации с |
тойчивый при высокой температуре, в присутствии |
||
|
|||
|
помощью ферментов, установление нуклеотидной |
Mn2+ действует как обратная транскриптаза, достра- |
|
|
последовательности ДНК, осуществление химиче- |
ивая цепь ДНК на РНК-матрице. После завершения |
|
|
ского синтеза ДНК определенной последовательно- |
реакции полученный гибридный дуплекс мРНК–ДНК |
|
|
сти; 2) клонирование и экспрессия генов в эукариоти- |
обрабатывают ферментом РНКазой и получают одно- |
|
|
ческих и прокариотических клетках. |
цепочечную ДНК, которую можно использовать в ка- |
|
Методы |
ВЫДЕЛЕНИЕ ДНК, ЕЕ АМПЛИФИКАЦИЯ И ОБРАБОТКА |
честве матрицы для создания двухцепочечной ДНК в |
|
МОДИФИЦИРУЮЩИМИ ФЕРМЕНТАМИ. Количество |
обычной ПЦР. Для неспецифической амплификации |
||
|
|||
|
ДНК в клетке очень мало’ (одна молекула ДНК на |
эукариотических мРНК используют участок полиаде- |
|
|
прокариотическую клетку и один или два набора хро- |
нилирования на 5'-конце мРНК: в качестве затравки |
|
|
мосом на эукариотическую клетку), тем не менее |
для синтеза кДНК на матрице мРНК служит олиго- |
|
|
ДНК из клеток можно выделить химическими мето- |
мер, состоящий из тимидиновых оснований (polyT). |
|
|
дами, выполнить ее анализа и использовать в даль- |
ЭКСПРЕССИЯ. К настоящему времени разработано |
|
|
нейших исследованиях. Как правило, для работы не |
несколько методов встраивания ДНК в ДНК другого |
|
|
применяют длинные цепи ДНК, а обрабатывают ее |
хозяина. После репликации в процессе клеточного |
|
|
специальными ферментами, которые могут расщеп- |
деления происходит транскрипция ДНК, а затем в ре- |
|
|
лять ДНК по определенным сайтам, осуществлять ее |
зультате трансляции образуются белки, среди кото- |
|
|
модификации, соединять фрагменты ДНК, а также |
рых и белок, закодированный в клонированном фраг- |
|
|
переводить ДНК в мРНК. Большинство эксперимен- |
менте ДНК. Совокупность этих процессов называется |
|
|
тов требует достаточно больших количеств ДНК: с |
экспрессией клонированного гена. В качестве кле- |
|
|
помощью автоматических синтезаторов можно полу- |
ток-хозяев, как правило, используют бактериальные |
|
|
чить лишь небольшие фрагменты ДНК, размером до |
клетки. Такой выбор обусловлен следующими |
|
|
100 п.н., поэтому особое значение приобретает ме- |
особенностями бактерий: 1) их геном представлен |
|
|
тод ПЦР, позволяющий амплифицировать фрагмен- |
всего лишь одной двухцепочечной молекулой ДНК; |
|
|
ты ДНК размером в тысячи пар нуклеотидов. Важное |
2) для переноса генетического материала можно |
|
|
применение метода ПЦР – рекомбинация между |
использовать фаги или плазмиды; 3) молекулярно- |
|
|
фрагментами ДНК из различных источников, напри- |
генетические основы организации многих бактерий |
|
|
мер между ДНК, выделенной из живого организма, и |
достаточно хорошо изучены; 4) бактерии быстро раз- |
|
|
химически синтезированной ДНК. |
множаются, и в биореакторах можно выращивать |
|
|
СВОЙСТВА ДНК И СЕКВЕНИРОВАНИЕ. Основные |
очень большие количества бактериальных клеток. |
|
|
характеристики ДНК: 1) температура плавления (ДНК |
Наряду с Escherichia coli для клонирования и экспрес- |
|
|
с бо’ льшим содержанием пар GC денатурирует при бо- |
сии чужеродной ДНК используют и другие бактерии, |
|
|
лее высокой температуре по сравнению с ДНК, обога- |
например Bacillus, Lactobacillus, Pseudomonas и |
|
|
щенной парами АТ); 2) молекулярная масса (этот па- |
Streptomyces. В последнее время разработаны мето- |
|
|
раметр устанавливают путем электрофоретического |
ды введения ДНК и в эукариотические организмы, |
|
|
разделения фрагментов ДНК в присутствии фрагмен- |
такие как дрожжи, грибы, а также в клетки растений |
|
|
тов ДНК известного размера); 3) последовательность |
и животных. |
|
|
нуклеотидов, которую устанавливают путем секвени- |
|
|
|
рования ДНК; 4) белковый продукт или функциональ- |
|
|
|
ный элемент, закодированный в исследуемой ДНК; |
|
|
|
5) взаиморасположение сайтов расщепления эндону- |
|
|
|
клеазами рестрикции (карта рестрикции). |
|
|
|
КЛОНИРОВАНИЕ. Методы, позволяющие выделить из |
|
|
|
генома прокариот участок, кодирующий определен- |
|
|
|
ный белок, также применимы и для эукариот, однако |
|
|
|
в этом случае необходимо учитывать наличие интро- |
|
|
|
нов в большинстве эукариотических ДНК. Зрелую |
|
|
|
(после сплайсинга) мРНК выделяют из клетки, а за- |
|
|
|
тем с помощью фермента обратной транскриптазы |
|
|
228 |
(ОТ) в реакции ОT-ПЦР синтезируют комплементар- |
|
|
ную цепь ДНК (кДНК). В лабораторной практике при- |
|
||
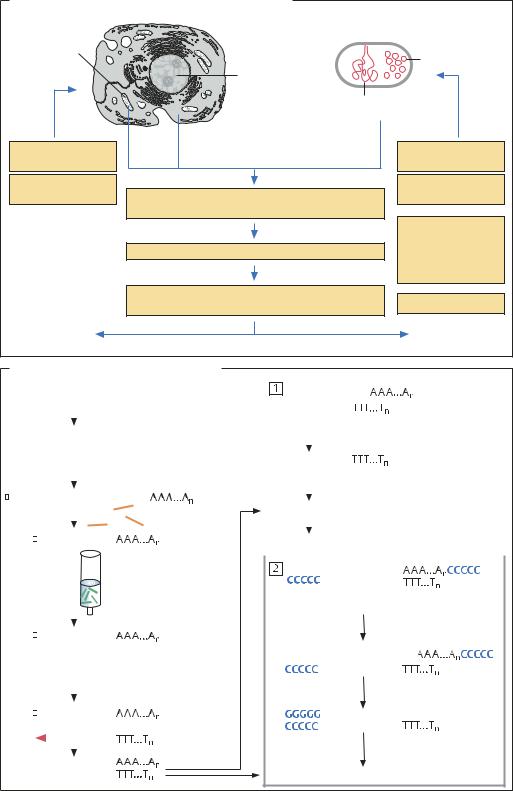
Основные стадии генно-инженерного эксперимента |
|
|
||
Эукариотическая клетка |
Прокариотическая клетка |
|||
Митохонд- |
|
|
|
|
риальная |
|
|
|
Плазмидная ДНК |
ДНК |
|
|
|
|
|
|
|
|
|
|
|
ДНК |
|
|
|
|
|
Нуклеоид, содержащий |
|
|
|
|
геномную ДНК |
|
|
Выде- |
Выделение мРНК |
Выде- |
|
Картирование генома |
ление |
и получение |
ление |
Картирование генома |
и секвенирование |
ДНК |
ее кДНК-копии |
ДНК |
и секвенирование |
Выключение |
|
|
|
Выключение |
определенных генов |
Обработка ДНК ферментами рестрикции |
|
определенных генов |
|
|
|
|
||
|
и/или амплификация фрагментов генома методом ПЦР |
|
||
|
|
|
|
Химический синтез |
|
Встраивание кДНК или фрагмента гена в векторную ДНК |
фрагментов ДНК |
||
|
и мечение праймеров |
|||
|
|
|
|
или ДНК-зондов |
|
Трансформация клеток-хозяев полученной векторной |
Секвенирование ДНК |
||
|
конструкцией, оптимизация условий экспрессии гена |
|||
Эукариотические |
|
|
|
Прокариотические |
клетки |
Трансформированные клетки синтезируют необходимый продукт |
клетки |
||
|
|
|||
Клонирование эукариотических генов |
|
|
||
|
Интрон |
|
Экзон |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Расщепление цепи |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Эукариотическая ДНК |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
РНК с помощью |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
Транскрипция |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
РНКазы H |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Синтез комплементарной |
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Первичный РНК-продукт |
|
|
|
|
|
цепи ДНК с помощью |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ДНК-полимеразы |
|||||||||||||
|
|
|
|
|
|
|
|
|
|
Кэпирование |
(содержащий интроны) |
|
|
|
|
|
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
и полиадени- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
лирование |
|
|
|
|
|
|
Обработка S1-нуклеазой |
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
для получения фрагмента |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ДНК с «тупыми» концами |
||||||||||||
|
|
|
|
|
|
|
|
|
|
Сплайсинг |
Зрелый |
|
|
|
|
|
|
|
|
|
|
|
Двухцепочечная кДНК |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Встраивание в вектор/трансформация клеток E. coli |
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
мРНК-продукт |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
(не содержащий |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Элюиро- |
|
|
|
Олиго-dТ |
интронов) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
Обработка терминальной трансферазой для создания |
||||||||||||||||||||||||||
|
|
|
|
вание |
|
|
|
целлюлоза |
||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
последовательности олиго-dС на концах |
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Расщепление цепи РНК |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Очищенный |
|
|
|
|
|
|
|
|
|
|
|
|
|
с помощью РНКазы Н |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
препарат |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Синтез цепи, |
зрелой мРНК |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
комплементарной |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
мРНК, с помощью |
|
|
|
|
|
|
|
|
|
|
|
|
|
Синтез комплементарной |
||||||||||
|
|
|
|
|
|
|
|
|
|
обратной транскриптазы |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
цепи ДНК с помощью |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
фрагмента Клёнова |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Зрелая мРНК |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Праймер |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Обработка S1-нуклеазой |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
для получения фрагмента |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ДНК с «тупыми» концами |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Встраивание в вектор/трансформация клеток E. coli |
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||
229
