

- •Содержание
- •Предисловие
- •Предисловие ко 2-му изданию
- •Введение
- •Этапы развития биотехнологии
- •Биотехнология сегодня
- •Биотехнологическое производство пищевых продуктов
- •Алкогольные напитки
- •Пивоварение
- •Ферментация в пищевой промышленности
- •Пищевые продукты и молочнокислое брожение
- •Этиловый спирт
- •1-Бутанол, ацетон
- •Уксусная кислота
- •Лимонная кислота
- •Молочная и глюконовая кислоты
- •Аминокислоты
- •L-Глутаминовая кислота
- •D,L-Метионин, L-лизин и L-треонин
- •Антибиотики
- •Антибиотики: источники, применение и механизмы действия
- •Антибиотики: получение. Устойчивость к антибиотикам
- •β-Лактамные антибиотики: промышленное получение
- •Гликопептидные, полиэфирные и нуклеозидные антибиотики
- •Аминогликозидные антибиотики
- •Тетрациклины, хиноны, хинолоны и другие ароматические антибиотики
- •Поликетидные антибиотики
- •Получение новых антибиотиков
- •Специальные продукты
- •Витамины
- •Нуклеозиды и нуклеотиды
- •Биодетергенты и биокосметика
- •Микробные полисахариды
- •Биоматериалы
- •Биотрансформация
- •Биотрансформация стероидов
- •Ферменты
- •Ферменты
- •Ферментативный катализ
- •Ферменты в клинических анализах
- •Тесты с помощью ферментов
- •Применение ферментов в промышленных технологиях
- •Ферменты в производстве моющих средств
- •Ферменты, расщепляющие крахмал
- •Ферментативное расщепление крахмала в промышленности
- •Ферментативное превращение сахаров
- •Утилизация целлюлозы и полиозы
- •Использование ферментов в целлюлозно-бумажной промышленности
- •Пектиназы
- •Ферменты в производстве молочных продуктов
- •Использование ферментов в хлебобулочной и мясоперерабатывающей промышленности
- •Ферменты в кожевенной и текстильной промышленности
- •Перспективы получения ферментов для промышленных технологий
- •Белковая инженерия
- •Пекарские и кормовые дрожжи
- •Пекарские и кормовые дрожжи
- •Белки и жиры из одноклеточных организмов
- •Аэробная очистка сточных вод
- •Анаэробная очистка сточных вод и переработка ила
- •Биологическая очистка газовых выбросов
- •Биологическая очистка почв
- •Микробиологическое выщелачивание руд и биокоррозия
- •Инсулин
- •Гормон роста и другие гормоны
- •Гемоглобин, сывороточный альбумин и лактоферрин
- •Факторы свертывания крови
- •Антикоагулянты и тромболитики
- •Ингибиторы ферментов
- •Иммунная система
- •Стволовые клетки
- •Тканевая инженерия
- •Интерфероны
- •Интерлейкины
- •Эритропоэтин и другие факторы роста
- •Другие белки, имеющие медицинское значение
- •Вакцины
- •Рекомбинантные вакцины
- •Антитела
- •Моноклональные антитела
- •Рекомбинантные и каталитические антитела
- •Методы иммуноанализа
- •Биосенсоры
- •Биотехнология в сельском хозяйстве
- •Животноводство
- •Перенос эмбрионов и клонирование животных
- •Картирование генов
- •Трансгенные животные
- •Генетическая ферма и ксенотрансплантация
- •Растениеводство
- •Культивирование растительных клеток: поверхностные культуры
- •Культивирование растительных клеток: суспензионные культуры
- •Трансгенные растения: методы получения
- •Трансгенные растения
- •Вирусы
- •Бактериофаги
- •Микроорганизмы
- •Бактерии
- •Некоторые бактерии, важные для биотехнологии
- •Грибы
- •Дрожжи
- •Усовершенствование штаммов микроорганизмов
- •Основы биотехнологических методов
- •Микроорганизмы: рост в искусственных условиях
- •Кинетика образования продуктов метаболизма и биомассы в культуре микроорганизмов
- •Технология ферментации
- •Промышленные процессы ферментации
- •Культивирование животных клеток
- •Биореакторы для культивирования животных клеток
- •Биореакторы с иммобилизованными ферментами и клетками
- •Очистка биотехнологических продуктов
- •Очистка биотехнологических продуктов: хроматографические методы
- •Экономические аспекты биотехнологического производства
- •Методы генетической инженерии
- •Структура ДНК
- •Функции ДНК
- •Эксперимент в генетической инженерии
- •Методы выделения ДНК
- •Ферменты, модифицирующие ДНК
- •ПЦР: лабораторная практика
- •ДНК: химический синтез и определение размера молекул
- •Секвенирование ДНК
- •Введение ДНК в живые клетки (трансформация)
- •Идентификация и клонирование генов
- •Экспрессия генов
- •Выключение генов
- •Геном прокариот
- •Геном эукариот
- •Геном человека
- •Функциональный анализ генома человека
- •ДНК-анализ
- •Белковые и ДНК-чипы
- •Маркерные группы
- •Тенденции развития
- •Генная терапия
- •Поиск биологически активных веществ
- •Протеомика
- •Обмен веществ
- •Метаболомика и метаболическая инженерия
- •Системная биология
- •«Белая» биотехнология
- •Сертификация биотехнологической продукции
- •Этические аспекты генетической инженерии
- •Патентование в биотехнологии
- •Биотехнология в разных странах
- •Биотехнология в разных странах
- •Литература
- •Источники иллюстраций
- •Указатель микроорганизмов
Биотехнология в сельском хозяйстве
182
Трансгенные растения
ВВЕДЕНИЕ. Изменение растений генно-инженерными методами преследует цель не только модификации их собственных веществ («растительного сырья»), но и синтеза ценных веществ в растениях. Например: 1) может происходить изменение аминокислотного состава, изменение содержания крахмала и масел (жиров), а в древесине – лигнина; 2) синтез чужеродных (для растения) веществ, таких как антитела или их фрагменты, вакцины, сывороточный альбумин человека или биополимеры.
МОДИФИКАЦИЯ СОБСТВЕННЫХ ВЕЩЕСТВ РАСТЕНИЯ.
Восполнение в растительных белках недостатка незаменимых аминокислот (преимущественно L-лизина и L-метионина) осуществляется несколькими вполне эффективными путями: а) экспрессией генов более выгодных в этом отношении белков из других растений; б) сайт-направленным мутагенезом генов этих белков для внедрения незаменимых аминокислот вместо заменимых; в) встраиванием генов, которые контролируют синтез необходимых белков (например, генов аспартаткиназы из Escherichia coli и синтазы дегидродипиколиновой кислоты из Corynebacterium для повышения выхода в синтезе L-лизина). Другой задачей является изменение содержания и состава крахмала. Активность АДФ-глюкозопирофосфорилазы, ключевого фермента в биосинтезе крахмала, регулируется во многих растениях аллостерически. Путем встраивания нерегулируемого гена из E. coli удается получить томаты с содержанием крахмала выше на ~20%. Отношение содержания линейной формы (амилозы) к разветвленной (амилопектину) определяет использование крахмала в пищевых или технических целях. Повышение уровня экспрессии гена glgB из E. coli, контролирующего образование α-1,6- связей между мономерами крахмала, в присутствии промотора гена синтазы гранулированного крахмала, у картофеля приводит к накоплению крахмала с повышенным почти на 25% содержанием амилопектина. Состав жирных кислот в масличных культурах также может быть изменен. Например, лауриновая кислота (C12), очень важный компонент для получения растворимых в холодной воде детергентов, в большом количестве содержится только в маслах тропических растений (в кокосовой и других пальмах) в форме триглицеринов. Путем встраивания гена специфичной к длине углеродной цепи ACP-эстеразы жирных кислот из лавра (Umbellularia california) (или иным способом) в высокопродуктивные селекционные линии рапса удается получить трансгенные растения, которые содержат в семенах до 50 моль % трилауроилглицерина. Содержание лигнина в древесине удается понизить путем модификации генов ферментов, участвующих в биосинтезе (стадии образования промежуточного метаболита – коричной кислоты).
ПОЛУЧЕНИЕ ЧУЖЕРОДНЫХ ДЛЯ РАСТЕНИЯ ВЕЩЕСТВ.
Большинство исследований в этой области проводят на растениях табака или Arabidopsis thaliana, которые особенно легко подвергаются трансформации. Так, возможна экспрессия гена человеческого сывороточного альбумина, а также получение IgG-антител, которые используются в профилактике кариеса против адгезина Streptococcus mutans. Общая концентрация целевых белков может достигать 1%. Дешевый метод получения антигенов в растениях открывает дорогу вакцинированию населения в развивающихся странах путем приема пищи. В модельных экспериментах удалось добиться того, что поверхностные антигены вируса гепатита B в табаке составили 0,01% содержания всех растворимых белков, и добавление порошка из трансгенного табака к рациону мышей приводило к развитию у них иммунного ответа. Потребление картофеля, содержащего энтеротоксин B из E. coli, также вызывает иммунный ответ у человека. Экспрессия оперона Alcaligenes eutrophus, состоящего из трех генов и ответственного за синтез полигидроксибутиратов, в хлоропластах Arabidopsis thaliana или рапса дает возможность получать этот способный к биодеградации пластик дешевым путем при фотосинтезе. С экономической точки зрения такой синтез этого биополимера все же нуждается в дальнейшем усовершенствовании.

Томатная паста
Из томатов повышенной сахаристости
Трансгенный рапс с измененным содержанием жирных кислот
мМЕ |
|
|
Дикий тип |
|
|
600 |
|
Трансгенное растение |
|
||
активность, |
|
|
с измененной активностью |
||
|
|
ацил-АСР-гидролазы |
|
||
|
|
|
|
||
Тиоэстеразная |
400 |
|
|
|
|
200 |
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
10:0 |
12:0 |
14:0 |
16:0 |
18:0 |
|
|
Длина цепи ацильных групп |
|
||
Полигидроксимасляная кислота в пластидах трансгенного рапса
|
|
|
|
|
|
|
|
|
|
|
|
CW |
= Клеточная стенка |
||||
|
|
|
|
|
|
|
|
|
|
|
|
P |
= Пластида |
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
PHB = Полигидроксимасляная |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
кислота |
|
|
||
|
|
|
|
|
|
|
PHB |
|
|
L |
= Липиды |
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
L |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
L |
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
P |
|
|
|
|
|
|
|
CW |
|
|
|
1 мкм CW |
|
|
|
1 мкм |
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Растения как биореакторы (примеры) |
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Растение |
Чужеродный/измененный |
Наблюдаемые |
|
|
|
Метод |
|
|||||||||
|
|
или инактивированный ген |
изменения свойств |
трансформации |
|
||||||||||||
|
Модификация собственных соединений растения |
|
|
|
|
|
|
|
|
||||||||
|
Рапс, соя |
Аспартокиназа (E. coli) и синтаза |
Повышенное содержание L-лизина |
Ti-плазмида |
|
||||||||||||
|
|
дигидродипиколиновой кислоты |
|
|
|
|
|
|
|
|
|||||||
|
|
(Corynebacterium) |
|
|
|
|
|
|
|
|
|||||||
|
Картофель, томат |
Крахмал-синтетаза |
Другой состав крахмала |
Антисмысловая |
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
конструкция |
|
|
Рапс |
Ацил-АСР-гидролаза |
Другой состав жирных кислот |
Ti-плазмида |
|
||||||||||||
|
Резуховидка |
γ-Токоферолметилаза |
Синтез витамина Е |
|
|
|
Ti-плазмида |
|
|||||||||
|
Ель |
Синтаза шикимовой кислоты |
Пониженное содержание лигнина |
Антисмысловая |
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
конструкция |
|
|
Экспрессия чужеродных соединений |
|
|
|
|
|
Выход продукта, г/кг |
|
|||||||||
|
Картофель |
Белок оболочки вируса гепатита В |
Иммунный ответ у мышей |
|
|
||||||||||||
|
|
Сывороточный альбумин человека |
Синтез сывороточного альбумина |
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
человека |
|
|
|
|
|
||
|
Табак |
Фрагмент IgG к адгезину вируса |
Синтез фрагментов антител |
10 |
|
||||||||||||
|
|
бешенства, Bacillus anthrax |
|
|
|
|
|
|
|
|
|||||||
|
Рапс, резуховидка |
Три гена (phb-оперон) |
Синтез полигидроксибутирата |
140 |
|
||||||||||||
|
|
из Alcaligenes eutrophus |
(ПГБ, или англ. РНВ) |
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
или полигидроксивалерата |
|
|
|||||
|
|
|
|
|
|
|
|
|
|
(ПГВ, или англ. PHV) |
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
Трансгенное растение |
|
|
Урожай |
|
|
Обработка |
|
|
До 140 г/кг |
|||||||
|
Оперон для PHV |
|
|
|
|
|
|
|
|
«фитополимеров» |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
183
микробиологии |
Вирусы |
|
ВВЕДЕНИЕ. Вирусы не обладают собственным обме- |
||
|
||
|
ном веществ. Репликация генетического материала |
|
|
вирусов – ДНК или РНК – происходит с помощью кле- |
|
|
ток-хозяев. Вне клетки-хозяина вирус представляет |
|
Основы |
собой нуклеиновую кислоту, одетую белковой оболоч- |
|
кой (капсидом). Такое состояние вируса называется |
||
нуклеокапсидом или вирионом. Вирусы могут инфици- |
||
ровать большинство живых организмов, однако они |
||
проявляют высокую специфичность по отношению к |
||
|
клеткам-хозяевам, выбирая определенные ткани или |
|
|
клетки в организме. Существует несколько способов |
|
|
классификации вирусов: по типам клеток-хозяев, по |
|
|
морфологическим признакам, по генетическому мате- |
|
|
риалу (ДНК или РНК) или по строению капсида. Виру- |
|
|
сы широко используются в биотехнологии для получе- |
|
|
ния многокомпонентных вакцин, а также для |
|
|
разработки различных векторов, например, для генной |
|
|
терапии или для экспрессии генов в культурах клеток. |
|
|
ВИРУСЫ В ЭКСПЕРИМЕНТАХ С ЖИВОТНЫМИ. Пер- |
|
|
вые эксперименты по клонированию животных кле- |
|
|
ток были проведены в 1979 г. с использованием |
|
|
вектора на основе вируса обезьян (SV40). Вирус про- |
|
|
никает в клетку, а затем его развитие протекает по |
|
|
литическому или лизогенному пути. Геном вируса |
|
|
(5,2 т.п.н.) содержит так называемые «ранние гены», |
|
|
кодирующие белки, необходимые для репликации |
|
|
ДНК, и «поздние гены», продукты которых участвуют |
|
|
в синтезе капсида. Векторы на основе SV40 содер- |
|
|
жат вирусные регуляторные элементы: точку начала |
|
|
репликации, промоторный участок, а также термина- |
|
|
тор транскрипции (сайт полиаденилирования). Для |
|
|
трансфекции клеток мышей используют конструкции |
|
|
на основе папилломавируса крупного рогатого скота |
|
|
(BPV). При инфекции они ведут себя как высококо- |
|
|
пийные плазмиды, и ДНК-копии передаются при кле- |
|
|
точном делении дочерним клеткам. Ослабленные ре- |
|
|
тровирусы, аденовирусы, а также вирус герпеса |
|
|
применяются для генной терапии. Геном ретровиру- |
|
|
сов (например, ВИЧ) представляет собой РНК. Рет- |
|
|
ровирусы инфицируют делящиеся клетки, при этом |
|
|
обратная транскриптаза, кодированная в вирусном |
|
|
геноме, обеспечивает синтез кДНК-копии вирусного |
|
|
РНК-генома. Эта кДНК встраивается в геном хозяина |
|
|
и использует его сильные промоторы для синтеза |
|
|
белков капсида и вирусной мРНК. В эксперименталь- |
|
|
ной генной терапии успешно используются ретрови- |
|
|
русы с дефектами репликации, однако размер ДНК, |
|
|
доставляемой с помощью вируса, невелик. В отличие |
|
|
от ретровирусов аденовирусы могут быть использо- |
|
|
ваны для упаковки крупных молекул ДНК (до |
|
|
28 т.п.н.). Аденовирусы инфицируют как делящиеся, |
|
|
так и неделящиеся клетки, однако их ДНК не встраи- |
|
184 |
вается в геном хозяина. Известен случай, когда ис- |
|
пользование аденовирусов вызвало неожиданную |
иммунную реакцию и привело к смерти 18-летнего пациента. С тех пор использование аденовирусов в медицине практически прекращено. В настоящее время изучается возможность использования векторов на основе вируса простого герпеса Нerpes simplex для генной терапии таких нервных расстройств, как болезни Паркинсона или Альцгеймера. Вирус Нerpes simplex обладает большим ДНК-геномом (152 т.п.н.), следовательно, в него можно встраивать крупные фрагменты ДНК.
ВИРУСЫ В ЭКСПЕРИМЕНТАХ С РАСТЕНИЯМИ.
Большинство вирусов растений имеют РНК-геном. Известны лишь две группы ДНК-содержащих вирусов, которые могут инфицировать высшие растения. У каулимовирусов спектр клеток-хозяев очень узкий: они поражают только представителей семейства крестоцветных – свеклу и некоторые сорта капусты. Каулимовирусы имеют очень маленький капсид, поэтому их собственный геном и чужеродная упакованная ДНК очень невелики. Геминивирусы инфицируют такие важные сельскохозяйственные культуры, как кукуруза и пшеница, поэтому их использование сопряжено с высокой степенью риска. Кроме того, при инфекции в геноме геминивирусов происходят множественные перестройки, в том числе и делеции ДНК, поэтому часто возникают проблемы с экспрессией встроенных фрагментов ДНК.
БАКУЛОВИРУСЫ. Эти вирусы заражают насекомых, но безопасны для позвоночных. В результате вирусной инфекции в клетке начинается синтез кристаллического белка полигедрина (полиэдрина), а синтез более половины белков клетки-хозяина подавляется. Промотор полигедрина используется при создании векторов для гетерологичной экспрессии в клеточной культуре Spodoptera (род бабочек). Премущество заключается в том, что посттрансляционное гликозилирование в такой системе аналогично таковому у позвоночных. В настоящее время перечисленные системы экспрессии используются только в лабораторных исследованиях.
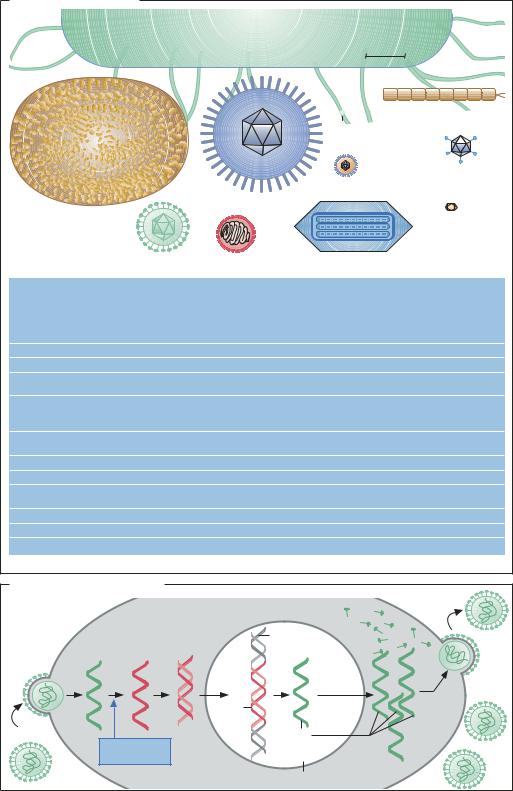
Многообразие вирусов
Вирус оспы
Для сравнения: клетка Escherichia coli
100 нм
 Вирус табачной мозаики Каулимовирус
Вирус табачной мозаики Каулимовирус 
Тогавирус |
Аденовирус |
|
Вирус |
||
|
||
герпеса |
|
|
|
Геминивирус |
|
ВИЧ |
Вирус гриппа |
Бакуловирус |
|
|
|
|
|
|
Вирус |
Организм-хозяин |
Заболевание, |
Оболочка |
Геном |
|
|
вызываемое |
|
|
|
|
вирусной инфекцией |
|
|
|
|
|
|
|
Вирус оспы |
Человек, |
Оспа |
Сложная оболочка |
Линейная ДНК, д |
|
домашние животные |
|
|
|
Вирус гепатита В |
Человек |
Гепатит В |
Полиэдрический капсид |
Кольцевая ДНК, д |
Тогавирус |
Человек |
Корь |
Полиэдрический капсид |
(+)-РНК, о |
Вирус герпеса |
Человек, птицы |
Опоясывающий |
Полиэдрический капсид, |
Линейная ДНК, д |
|
|
лишай и др. |
наружная оболочка |
|
ВИЧ (вирус |
Человек, приматы |
СПИД (синдром |
Сферическая оболочка |
2 × (+)-РНК, о |
иммунодефицита |
|
приобретенного |
|
|
человека) |
|
иммунодефицита) |
|
|
Вирус гриппа |
Человек, |
Грипп |
Спиралевидный, |
(–)-РНК, |
|
млекопитающие |
|
с внешней оболочкой |
сегментированная |
Аденовирус |
Человек |
Простуда |
Полиэдрический капсид |
Линейная ДНК, д |
Папилломавирус |
Скот |
Бородавки |
Полиэдрический капсид |
Кольцевая ДНК, д |
Вирус табачной |
Табак |
|
Полиэдрический капсид |
РНК, о |
мозаики |
|
|
|
|
Каулимовирус |
Капуста |
|
Полиэдрический капсид |
Кольцевая ДНК, о |
Геминивирус |
Двудольные |
|
Двойной полиэдр |
Кольцевая ДНК, о |
Бакуловирус |
Насекомые |
|
Полиэдрический капсид |
Кольцевая ДНК, д |
|
о – одноцепочечная, д – двухцепочечная, + – «смысловая» РНК, – - «антисмысловая» РНК |
|
||||
Цикл развития ретровируса |
|
|
|
||
|
|
|
|
Отпочковывание |
|
|
|
Одно- |
ДНК |
Белок капсида |
|
|
|
|
|
||
|
РНК |
цепо- |
(двух- |
Хромосома |
|
|
чечная |
цепо- |
|
||
|
хозяина |
|
|||
|
|
ДНК |
чечная) |
|
|
|
|
|
|
||
|
|
|
Про- |
|
Упаковка |
|
|
|
|
|
|
|
|
|
вирус |
|
|
Инфекция |
|
|
|
РНК |
|
|
Обратная |
|
вирусного |
|
|
Ретро- |
транскриптаза |
|
генома |
|
|
|
|
Клетка хозяина |
Клеточное ядро |
|
|
вирус |
|
|
185 |
||
|
|
|
|
|
|