


Г Л А В А 1 |
ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Рис. 1.7 Стимулы, используемые для иллюстрации наличия слепого пятна и феномена «запол нения». Закройте левый глаз. Зафиксируйте взгляд правого глаза на кресте и меняйте дистан цию просмотра до тех пор, пока точка (а) или прорезь в линии (b) не упадeт на ваше слепое пят но. Обратите внимание на ощущения в каждом случае.
быть проиллюстрирована рис. 1.7 (b), предназначенным для исследования сле пого пятна: закройте левый глаз, а взгляд правого зафиксируйте на кресте, за тем меняйте расстояние просмотра до тех пор, пока прорезь в линии не исчез нет, то есть пока она не попадет на слепое пятно, — вместо прорези вы будете видеть непрерывную линию, поскольку теперь это наиболее вероятный зри тельный стимул. Если вы хотите выполнить эти упражнения, закрыв левый глаз, то просто переверните рисунок вверх ногами, чтобы поймать слепое пятно на другой стороне поля зрения.
Феномен заполнения углубляет наше понимание механизмов работы зри тельной системы: сигналы, переданные ганглиозным клеткам, несут информа цию только о локальных изменениях в зрительных стимулах, причем дейст венна лишь информация о пространственных и временных переходах (то есть о границах), передаваемая в мозг. Восприятием этот код интерпретируется пу тем анализа характера изменений и заполняется соответствующим однород ным восприятием до поступления сигнала о новом переходе, что обеспечивает гигантскую экономию в полосе пропускания передаваемых сигналов. Можно даже говорить о некоем подобии группового кодирования (сжатия), которое иногда применяется в технологии цифровых изображений.
1.3 ОБРАБОТКА ЗРИТЕЛЬНОГО СИГНАЛА
Процесс нейронной обработки зрительной информации на уровне сетчатки сложен, а на дальнейших стадиях он становится значительно (если не бесконеч но) более сложным. Данный раздел — это краткий обзор путей, по которым идет зрительная информация, и имеет смысл начать его с общей карты этих путей.
Оптическое изображение на сетчатке вначале преобразуется в химические и электрические сигналы фоторецепторов, а затем эти сигналы обрабатывают ся клетками нейронной сети сетчатки (горизонтальными, биполярными, амак риновыми и ганглиозными клетками, описанными выше). Аксоны ганглиоз ных клеток формируют зрительный нерв, который идет к т.н. латеральному
40
Г Л А В А 1 |
|
|
|
|
|
ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
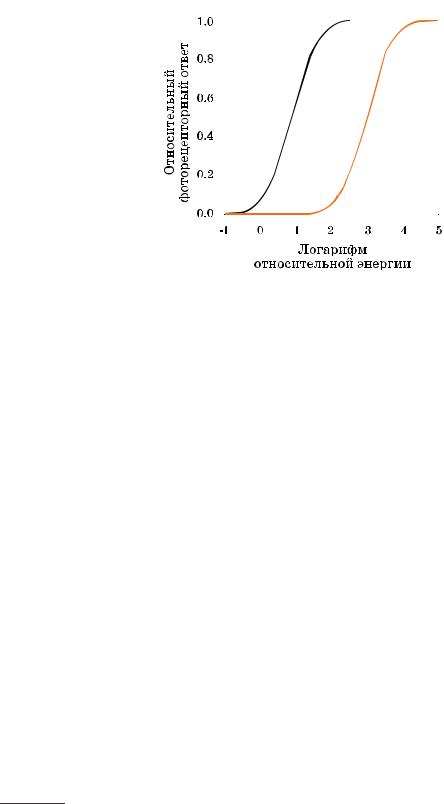
Рис. 1.8 Относительные энергетические ответы палочковых и колбочковых фоторецепторов.
коленчатому телу (lateral geniculate nucleus — LGN) в таламусе1. Клетки LGN, получив входной сигнал от ганглиозных клеток, отсылают его в т.н. первую зрительную область (V1) затылочных долей коры головного мозга, в которых обработка информации становится невероятно сложной. В коре головного моз га было найдено почти 30 зрительных областей, названные затем как V2, V3, V4, MT и т.д. Сигналы от этих областей отсылаются к некоторым другим облас тям и обратно. Корковая обработка включает в себя множество стадий: пря мую, обратную и латеральную обработки; где то в недрах этой сети и формиру ется наше окончательное восприятие. Кое какие детали этих процессов описа ны в следующих параграфах.
Свет, воздействующий на сетчатку, абсорбируется фотопигментом различ ных фоторецепторов (в палочках, в частности, это родопсин). После поглоще ния фотона родопсин меняет свою структуру, происходит цепная химическая реакция, итогом которой является закрытие ионных каналов в клеточной стен ке, продуцирующее электрический сигнал, основанный на разности относи тельных концентраций различных ионов (к примеру, натриевых и карбонат ных) вне и внутри клетки. Аналогичные процессы протекают в колбочках, ро допсин которых состоит из опсина и ретиналя: колбочки всех трех типов име ют сходную фотопигментную структуру, но у колбочковых опсинов («конопси нов») немного разные молекулярные формулы, что ведет к отличиям в спек тральной чувствительности колбочек. Каждый тип колбочек (L, M или S) со держит свою форму «конопсина».
На рис. 1.8 показаны относительные ответы фоторецепторов как функция световой экспозиции сетчатки. Любопытно, что характеристики этих функций подобны характеристикам многих устройств ввода и вывода изображений.
1 |
Таламус — подкорковый отдел головного мозга. Латеральное коленчатое тело — подструк |
|
|
тура таламуса. — Прим. пер. |
|
41
Г Л А В А 1 |
ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА |
В нижней части графика виден порог, ниже которого рецепторы не отвечают. Затем следует более или менее линейный участок кривых, и за ним — финаль ный участок — зона насыщения (верхний конец кривых). Приведенные кри вые описывают рецепторный фототок и демонстрируют самую первую стадию зрительной обработки. В дальнейшем сигналы обрабатываются нейронами сет чатки и синапсами. Результатом этой обработки является преобразованный сигнал, который ганглиозные клетки отправляют по зрительному нерву в мозг.
Рецептивные поля
По ряду причин (таких, как необходимость подавления шумов и обеспече ние высокой скорости передачи) амплитудно модулированные сигналы фото рецепторов конвертируются ганглиозными клетками и вышестоящими отде лами мозга в частотно модулированное представление. Сегодня несомненно то, что величина сигнала от возбужденной нервной клетки представлена количе ством пиков напряжения в секунду, а не разностью потенциалов по обе стороны клеточной стенки.
Чтобы получить представление о физиологических свойствах нервных кле ток сетчатки, необходимо познакомиться с концецпией т.н. рецептивных полей.
Рецептивное поле фоторецептора — это графическое представление неко ей области поля зрения, на которую отвечает данная клетка. На схеме рецеп тивного поля обычно указывают характер ответа различных областей этого поля (к примеру, положительный ответ, отрицательный ответ, спектральный сдвиг). Фактически рецептивное поле фоторецептора — это маленькая круглая область, указывающая размер и местоположение персональной чувствитель ности данного рецептора (в поле зрения).
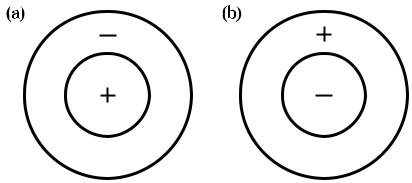
На рис. 1.9 показаны некоторые варианты рецептивных полей ганглиозных клеток, иллюстрирующие антагонизм центра и окружения, характерный для данного уровня зрительной обработки. Рецептивное поле на рис. 1.9 (а) иллю стрирует положительный центральный ответ на положительный входной сиг нал от единичной колбочки: положительный ответ окружен т.н. отрицатель ным ответом окружения, который, как правило, запускается отрицательным входным сигналом от соседних колбочек. Таким образом, ответ от данной ганг
Рис. 1.9 Типичные антагонистичные рецептивные поля «центр — окружение»: (а) — централь ное; (b) периферийное.
42
Г Л А В А 1 |
|
|
|
ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
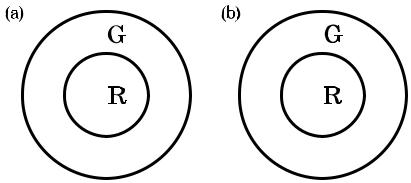
Рис. 1.10 Примеры красно зеленых (а) и зелено красных (b) спектрально и пространственно антагонистичных рецептивных полей.
лиозной клетки задается совокупностью положительных и отрицательных входных сигналов от множества колбочек, в результате чего ганглиозная клет ка не просто отвечает на точечный свет, но выступает в роли т.н. краевого дат чика (фактически — датчика «пятен»). Читатели, знакомые с цифровой обра боткой изображений, могут провести аналогию между ответом ганглиозной клетки и результатом операции свертки с заданным ядром, применяемой для обнаружения края.
На рис. 1.9 (b) показано, что ответы с противоположными полярностями рав новероятны: ответ, показанный на рис. 1.9 (а), рассматривают как ответ цен тральной ганглиозной клетки, тогда как ответ, показанный на рис. 1.9 (b), — на зывают ответом периферийной ганглиозной клетки. Зачастую центральные и периферийные клетки имеют одну и ту же пространственную локализацию, питаемую одними и теми же фоторецепторами, в результате чего повышается динамический диапазон системы.
Отметим, что ганглиозные клетки, показанные на рис. 1.9, не будут выда вать ответ на однородные поля (то есть тогда, когда положительные и отрица тельные сигналы сбалансированы). Сказанное выше иллюстрирует один из ас пектов компрессии изображения, выполняемого сетчаткой, благодаря которо му головной мозг не перегружается избыточной информацией — передаются данные лишь об изменениях в окружающем мире. Отметим, что пространст венная обработка информации в зрительной системе — это базис влияния фона на цветовое восприятие.
Рис. 1.9 иллюстрирует пространственную оппонентность ответов ганглиоз ных клеток, а на рис. 1.10 показано, что в дополнение к пространственной оп понентности к ответам ганглиозных клеток часто прибавляется спектральная оппонентность. На рис. 1.10 (а) показан красно зеленый оппонентный ответ в центре, питаемый положительным сигналом от L колбочки, при этом окру жение питаемо отрицательным сигналом от M колбочек. На рис. 1.10 (b) пока зана периферийная версия данной клетки. Таким образом, прежде чем зри тельная информация покинет сетчатку, происходит ее обработка, радикально влияющая на цветовое восприятие. Рисунки 1.9 и 1.10 демонстрируют типич ные рецептивные поля ганглиозных клеток, и хотя существует множество
43
Г Л А В А 1 |
ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА |
иных типов и вариантов ответа ганглиозных клеток, все они работают по сход ным принципам.
На пути к первичной зрительной коре сигналы проходят через латеральное коленчатое тело (LGN). Аксоны ганглиозных клеток заканчиваются в LGN, создавая с его клетками синапсы, что обеспечивает непосредственный («один в один») обмен информацией между ганглиозными и LGN клетками. Таким об разом, рецептивные поля LGN клеток идентичны таковым у ганглиозных кле ток. Возможно, LGN действует только как ретранслятор сигналов. Однако ве роятно и то, что LGN все же выполняет некую зрительную функцию, принимая на себя нейронные проекции из коры, которые, в свою очередь, могут отвечать за некоторые виды коммутации или за адаптационный механизм обратной свя зи. Аксоны LGN клеток подходят к первой зрительной области коры головного мозга (V1).
Обработка в V1;области
ВV1 области коры головного мозга кодирование зрительной информации существенно усложняется: подобно тому как исходящие сигналы от различ ных фоторецепторов сравниваются и объединяются для реализации ответов ганглиозной клетки, выходящие сигналы от LGN клеток сравниваются и объе диняются для производства кортикальных (корковых) ответов. Когда сигналы попадают в цепь кортикальной обработки, эта обработка повторяется несколь ко раз с резко возрастающим уровнем сложности, и в итоге рецептивные поля начинают терять свое значение.
ВV1 встречаются клетки, которые отвечают лишь на стимулы определенно го вида, например:
—края и полоски различной ориентации;
—вход от одного глаза, второго и обоих глаз;
—различные пространственные частоты;
—различные временные частоты;
—специфические пространственные местоположения;
—комбинации перечисленных стимулов.
Кроме того, обнаруживаются клетки, которые линейно комбинируют сиг налы от клеток LGN (и от прочих клеток с нелинейным суммированием).
Все варианты ответов необходимы для восприятия размера, формы, место положения, движения, интенсивности и цвета. Учитывая сложность корти кальных ответов в клетках V1, нетрудно представить себе, до какой степени сложны зрительные ответы, получаемые из сети, состоящей примерно из 30 зрительных областей.
На рис. 1.11 показана схема малой доли связей между различными извест ными кортикальными областями. Отметим, что рис. 1.11 демонстрирует связи именно между областями, но не между клетками: при этом существует пример но 109 нейронов коры, выполняющих персональные зрительные функции, деятельность которых очень сложно объяснить простыми словами. Однако функция отдельной клетки, как правило, не имеет значения, поскольку раз
44