

Глава 3. Строение глазного яблока

Точка сращения
Щелевой
контакт Базальная
поверхность пигментированного
слоя
Апикальная
поверхность
Базальная
поверхность беспигментного
слоя
Плотный контакт
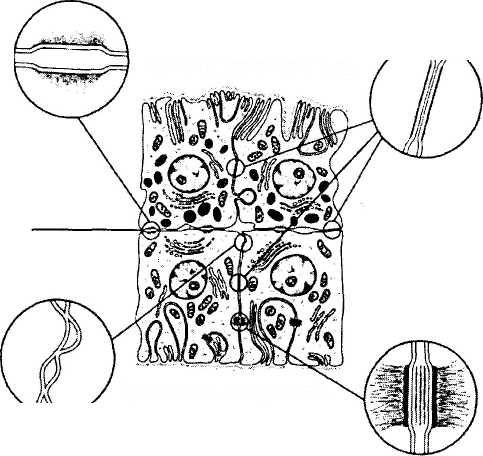
Рис. 3.8.35. Типы межклеточных контактов, обнаруживаемых между клетками пигментного эпителия ресничного тела (по Raviola et al., 1977)

Рис. 3.8.36. Электроннограмма эпителия ресничного тела:
/—внутренняя пограничная мембрана; 2—митохондрии; 3— гладкий эндоплазматический ретикулум; 4 — аппарат Гольджи; 5 — цитоплазматические отростки, распространяющиеся в подлежащую строму пигментных эпителиальных клеток; 6 — базальная мембрана пигментных эпителиальных клеток; 7 — меланино-вые гранулы пигментированного слоя эпителия
Кроме указанных контактов между двумя эпителиальными клеточными слоями обнаруживается и третий компонент, участвующий в межклеточном соединении — межклеточная цементирующая субстанция [317]. Она расположена внеклеточно между апикальными поверхностями клеточных мембран и имеет вид сплошной линии.
Беспигментный слой пигментного эпителия и внутренняя базальная мембрана. Внут-ренная базальная мембрана ресничного тела располагается с базальной (витреальной) стороны беспигментных клеток эпителия. Она связана с внутренней пограничной мембраной сетчатки сзади и радужки — спереди.
У новорожденных внутренняя базальная мембрана имеет типичное строение базальной мембраны. Состоит она из зернистого слоя толщиной 30 нм и прозрачного слоя (lamina lucida), толщина которого равна 50 нм. Состоит она из коллагеновых волокон I, III и IV типов, а также ламинина.
В возрасте 3 лет базальная мембрана начинает утолщаться и становится многослойной.
По направлению базальной мембраны ба-зальные поверхности беспигментных клеток образуют многочисленные складки и отростки, которые оплетаются волокнами мембраны и коллагеновыми волокнами основания стекловидного тела.
Беспигментные эпителиоциты имеют кубическую форму в отростчатой части ресничного тела (ширина 12—15 мкм и высота 10—15). В плоской части ресничного тела они цилиндрические (ширина 6—9 мкм и высота 30 мкм). Наиболее высокие клетки лежат вблизи зубчатой линии, возможно, в результате их тракции стекловидным телом. Многими исследователями отмечено увеличение высоты клеток с возрастом.
Между лежащими по соседству беспигментными эпителиальными клетками обнаруживаются щелевидные пространства различной ширины [317]. Эти пространства заполнены прозрачным внеклеточным муцинозным материалом, положительно окрашивающимся при проведении гистохимических реакций, направленных на выявление кислых гликозаминогли-канов.
Цитоплазма эпителиоцитов насыщена органоидами, что указывает на их высокую секреторную функцию. Особенно много в цитоплазме митохондрий [443]. Митохондрии являются основным источником энергии для обеспечения активного транспорта электролитов и молекул небольшого размера через цитоплазму эпителиальных клеток в процессе формирования камерной влаги.
В эпителиальных беспигментных клетках также хорошо развит шероховатый и гладкий эндоплазматический ретикулум [317, 496]. Цитоплазма эпителиоцитов содержит большое ко-
Сосуды и сосудистая оболочка глазного яблока
305
 личество
промежуточных микрофиламентов и
филаментов
актина. Необходимо отметить, что
микрофиламенты
положительно окрашиваются при
выявлении виментина и цитокератана.
личество
промежуточных микрофиламентов и
филаментов
актина. Необходимо отметить, что
микрофиламенты
положительно окрашиваются при
выявлении виментина и цитокератана.
Боковые поверхности беспигментных клеток образуют многочисленные складки цитоплазма-тической мембраны, погруженные в цитоплазму соседних клеток (интердигитации). Складки значительно увеличивают площадь контакта между клетками, что наиболее выражено в передней части ресничных отростков [154, 795, 872, 875, 890, 1125].
Апикальные поверхности беспигментных эпи-телиоцитов ресничных отростков при макроскопическом исследовании белые, что связано с высоким содержанием в их цитоплазме гликогена.
На переднем склоне ресничных гребешков по мере приближения к радужной оболочке в части эпителиальных клеток появляются зерна меланина. Степень пигментации постепенно нарастает, пока они не переходят в слой пигментного эпителия корня радужки.
Хотя внутренний слой ресничного эпителия и называется непигментированным, с возрастом возможно появление в их цитоплазме зерен меланина и липофусцина.
Большое значение в функционировании эпителия ресничного тела имеют межклеточные контакты, расположенные как между беспигментными клетками, так и между беспигментными и пигментированными клетками [872, 874, 875] (рис. 3.8.34, 3.8.35). Наиболее важным в функциональном смысле типом контакта является так называемая лента замыкания, располагающаяся на боковых поверхностях беспигментных клеток в апикальной их части. Встречаются также контакты типа «кепки», соединяющие боковые поверхности пигментированных и реже беспигментных клеток, щелевые контакты и десмосомы. Особое внимание в настоящее время уделяется изучению щелевых контактов. Предполагают, что эти контакты обеспечивают существование так называемого «электрического» взаимодействия эпителиальных клеток между собой. Именно подобный тип взаимодействия позволяет эпителиальному пласту функционировать как синцитий и, возможно, обеспечивает координацию секреторной деятельности всего эпителиального пласта.
Наличие перечисленных контактов между клетками, особенно ленты замыкания, делает практически невозможным проникновение между клетками высокомолекулярных веществ путем диффузии [1016].
Как неоднократно указывалось выше, беспигментный слой эпителиальных клеток ресничного тела является основным структурным элементом, обеспечивающим секрецию камерной влаги. Вышеприведенные светооптические, ультраструктурные особенности клеток, набор органоидов и плотность межклеточных контактов способствуют этому процессу. Из кровяно-
го русла капилляров ресничных отростков в переднюю камеру ионы и вещества небольшой молекулярной массы проходят благодаря функционированию энергозависимой транспортной системы [205, 206]. В соответствии с Cole [205] непигментированные клетки ресничного эпителия выборочно поглощают ионы натрия из стромы ресничного тела и транспортируют их через межклеточные пространства. Гиперосмо-тичность, возникающая в межклеточных пространствах в результате этого процесса, приводит к осмотическому потоку воды первоначально из стромы в межклеточные пространства, а затем в камерную влагу. О правильности подобного предположения свидетельствует обнаружение в мембранах непигментированных клеток ресничного эпителия ряда ферментов, участвующих в этом процессе, в частности активности Na+/K+-ATO-a3bi, угольной ангидразы [154, 327, 674].
Подтверждением тому являются и данные, относительно того, что применение средств, ин-гибирующих активность №+/К+-АТФ-азы, уменьшает секрецию камерной влаги. Прохождение ионов хлорида, бикарбоната и калия также обеспечивается активным транспортом. Ряд веществ проходит через эпителий пассивно, благодаря перепаду градиента концентрации. Это относится к сахарам и аминокислотам [241].
Секреция бикарбонатов снижается при применении ингибиторов угольной ангидразы (кар-боангидраза), что и используют при глаукоме для снижения секреции камерной влаги. Такими же свойствами обладают бета-блокаторы.
Беспигментные эпителиоциты ресничного тела, помимо секреции камерной влаги, участвуют еще во многих процессах.
Они синтезируют компоненты стекловидного тела — коллагены различного типа и глюку-роновую кислоту [124, 133, 716]. Участвуют эти клетки и в синтезе неколлагеновых белков — оптицина, фибулина-1 и нидогена-1, играющих немаловажную роль в стабилизации геля стекловидного тела и организации базальных мембран [717, 879].
Помимо основных секретирующих функций беспигментный эпителий принимает участие во многих репаративных и адаптационных процессах. Так, совсем недавно установлено, что он способен ингибировать пролиферативную активность и функции Т-лимфоцитов [1213], регулируя тем самым иммунные механизмы внутри глазного яблока.
Пигментированный эпителий и наружная базальная мембрана (рис. 3.8.32—3.8.36). Как было указано выше, пигментный слой эпителия ресничного тела развивается из наружного слоя зрительного бокала. По направлению к задним отделам глаза он переходит в пигментный эпителий сетчатой оболочки, а по направлению кпереди в передний слой пигментного эпителия радужки.
306