

Глава 3. Строение глазного яблока
 честве,
чем любые другие клетки глаза за
исключением
рецепторных клеток. Обнаруживаются
хорошо развитый гранулярный и
аграну-лярный эндоплазматический
ретикулум, многочисленные
свободные рибосомы. Вблизи ядра четко
виден аппарат Гольджи. Центриоли с
ресничками
располагаются в апикальной части
клеток.
В большом количестве определяются
лизосомы.
Отличительной чертой эндотелиаль-ных
клеток является наличие многочисленных
пиноцитозных
пузырьков, связанных с цито-плазматической
мембраной (рис. 3.2.11). Экспериментальными
исследованиями с использованием
радиоактивной метки показано быстрое
перемещение
этих пузырьков через цитоплазму
в сторону десцеметовой мембраны.
Иммуно-гистохимически
в цитоплазме эндотелиальных клеток
выявлены основные гликозаминоглика-ны
роговицы — хондроитин-6-сульфат,
хондрои-тин-4-сульфат,
гепаран-сульфат.
честве,
чем любые другие клетки глаза за
исключением
рецепторных клеток. Обнаруживаются
хорошо развитый гранулярный и
аграну-лярный эндоплазматический
ретикулум, многочисленные
свободные рибосомы. Вблизи ядра четко
виден аппарат Гольджи. Центриоли с
ресничками
располагаются в апикальной части
клеток.
В большом количестве определяются
лизосомы.
Отличительной чертой эндотелиаль-ных
клеток является наличие многочисленных
пиноцитозных
пузырьков, связанных с цито-плазматической
мембраной (рис. 3.2.11). Экспериментальными
исследованиями с использованием
радиоактивной метки показано быстрое
перемещение
этих пузырьков через цитоплазму
в сторону десцеметовой мембраны.
Иммуно-гистохимически
в цитоплазме эндотелиальных клеток
выявлены основные гликозаминоглика-ны
роговицы — хондроитин-6-сульфат,
хондрои-тин-4-сульфат,
гепаран-сульфат.
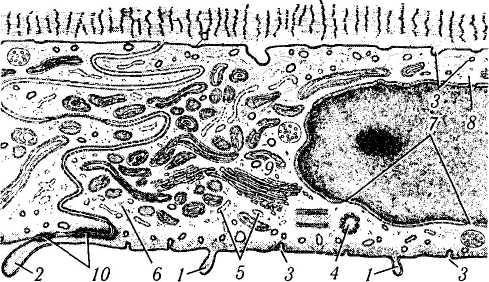
Рис. 3.2.11. Схематическое изображение ультраструктурной организации клеток эндотелия роговой оболочки:
/— микроворсинки; 2 — краевые выпячивания цитоплазмы в переднюю камеру глаза в местах межклеточных контактов; 3 — пиноцитозные пузырьки; 4 — центриоли; 5 — шероховатый эндоплазматический ретикулум; 6 — рибосомы; 7—ядерные поры; 8 — внутрицитоплазматические филаменты; 9 — аппарат Гольджи; 10 — межклеточные контакты различного типа
Необходимо остановиться и на основных физиологических функциях эндотелия роговицы. Одной из них является обеспечение клеток стромы питательными веществами. Процесс транспорта питательных веществ обеспечивается или диффузией между эндотелиоцитами, или активным переносом через содержимое клетки в направлении стромы.
Эндотелий играет главную роль в поддержании прозрачности роговицы путем активной регуляции содержания в строме воды. Эту функцию он выполняет, используя два механизма. Во-первых, он является активным барьером для солей и ряда метаболитов, проникновение которых в строму приводит к отеку последней. Во-вторых, он активно снижает осмотическое давление стромы наличием так называемого би-карбонатного насоса, возвращающего ионы из
стромы назад в камерную влагу [318, 711, 746, 918, 1204].
Кровоснабжение и лимфатическое дренирование роговицы обеспечиваются конъюнкти-вальными, эписклеральными и склеральными сосудами, являющимися ветвями передних ресничных артерий.
Нервы роговицы. Эпителий роговицы относится к наиболее интенсивно иннервируемым структурам организма человека. Чувствительная иннервация роговицы в 300—600 раз выше, чем иннервация кожи. Площадь эпителиального пласта, равная 0,01 мм2, содержит до 100 нервных окончаний [931]. На 2,1 млн ба-зальных клеток эпителия роговицы приходится до 1,4 млн нервных окончаний.
Сенсорная иннервация обеспечивается, в первую очередь, ветвями глазного нерва (ветвь тройничного нерва) [30, 878]. Главный источник иннервации роговицы — длинные ресничные нервы, являющиеся ветвями тройничного нерва.
Задний длинный ресничный нерв входит в склеру у заднего полюса и распространяется кпереди в супрахориоидальном пространстве. Различаются три уровня проникновения нервных окончаний в роговицу: склеральный, эпи-склеральный и конъюнктивальный [1222].
Около 80 нервных стволов проникают в ткань склеры вблизи лимба и, распространившись на 1—2 мм, теряют свои миелиновые оболочки. Эти волокна, покинув склеру, распределяются в средней трети стромы, делясь при этом ди- и трихотомически. Формируется в результате этого прекорнеальное сплетение. По мере продвижения к центральным участкам роговицы количество аксонов увеличивается за счет их последующего деления (рис. 3.2.12). При прохождении в строме роговой оболочки немиелинизованные нервные волокна располагаются параллельно коллагеновым пластинам. Отдельные нервные веточки подходят к керато-цитам и вдавливаются в их цитоплазматичес-кую оболочку [762, 763]. Окружают нервные стволы шванновские клетки и аморфный материал. Содержат аксоны многочисленные митохондрии, частицы гликогена и микропузырьки. Диаметр аксонов нервных волокон роговицы колеблется от 1 до 5 мкм.
В эпителиальный пласт из стромы нервы проникают через отверстия в боуменовой оболочке и образуют подэпителиальное сплетение [705, 971, 762, 763]. Иннервируются все эпителиальные клетки вплоть до поверхностных двух слоев, в которых нервные окончания имеют вид бусинок, колб Краузе, пластинок, лопаточек и др. [28, 29, 496, 762, 763, 878]. Концевые колбы Краузе, обеспечивающие температурную чувствительность, обнаруживаются лишь в области лимба. Некоторые сплетения нервных волокон контактируют с клетками Ларгенганса [971, 762, 763]. Иннервации дес-
Роговая оболочка и склера
181

/
КГ Г
V
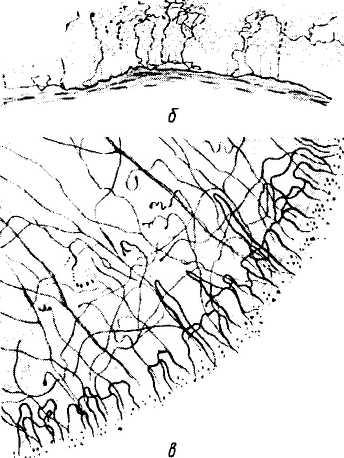
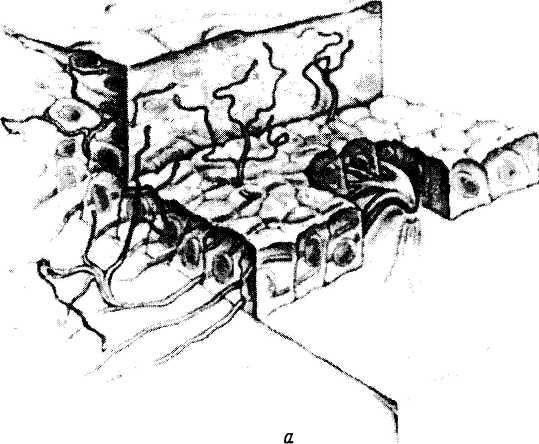
а — трехмерное изображение прохождения и распределения нервных волокон в роговой оболочке; б—поперечный срез роговицы. Распределение нервных волокон и нервных окончаний в переднем эпителии роговой оболочки; в — плоскостной препарат. Поверхностное краевое нервное сплетение
цеметовой оболочки и эндотелия не выявляются [931].
Ультраструктурные особенности нервов роговой оболочки позволяют некоторым авторам предполагать наличие пептидэргической иннервации как кератоцитов, так и эпителиальных клеток [762, 763].
Время регенерации нервных волокон роговицы длится около трех месяцев. Начинается регенерация нервов с периферии роговицы по направлению к центру. Помимо чувствительной иннервации, роговица обеспечена и вегетативной. Вегетативные волокна исходят из трех ганглиев. Это тройничный, ресничный и верхний шейный ганглии. Основным доказательством наличия вегетативной иннервации роговой оболочки является обнаружение отхождения нервных веточек от нервов лимбальных сосудов [1, 2, 28], а также эспериментальные исследования по перерезке нервных стволов, отходящих от вегетативных узлов, или после «раздражения» последних. Вегетативная иннервация обеспечивает трофику роговицы. Денервация роговой оболочки в эксперименте путем перерезки нервных стволов, входящих в глазное яблоко вблизи зрительного нерва, приводит к развитию дистрофических процессов, напоминающих нейропаралитический кератит у человека [16,17, 30]. Аналогичного характера дистрофические процессы роговой оболочки и структур переднего отдела глаза наблюдаются и после проведения циркляжа силиконовой лентой, которая передавливает ресничные нервы [12].
О значении иннервации говорит и то, что одним из необходимых основных условий диф-
ференциации эпителиальных и стромальных компонентов роговицы после травмы или кератопластики является реиннервация роговой оболочки [6, 30].
Старение роговой оболочки. Старение является естественным процессом у многоклеточных животных, приводящим к нарушению структуры и функции тканей и органов [923]. У человека признаки старения проявляются как функция времени. Выражаются они в нарушении дифференциации клеток, а их причиной являются биологические изменения, заложенные генетически или возникающие под влиянием на организм внешней среды.
Процесс старения ткани можно разделить на старение длительно существующих белков, старение делящихся клеток и старение неделя-щихся клеток [175].
Делящиеся клетки характеризуются тем, что их популяция поддерживается равновесием двух разнонаправленных процессов — скоростью размножения клеток и скоростью их гибели. Наиболее типичным примером такой популяции являются клетки переднего эпителия роговой оболочки. В роговичном эпителии полная смена дифференцированных эпителиальных клеток происходит за 5—7 дней [647, 695, 698]. Некоторые типы клеток способны к интенсивной пролиферации только после воздействия на них каких-либо внешних факторов. К подобным клеткам можно отнести кератоциты стромы роговицы [1115]. Клетки эндотелия роговицы также способны к делению, но деление эндотели-альных клеток у человека происходит исключительно редко [744, 977, 1087, 1127]. К неде-
182