

Инактивация X хромосомы у млекопитающих
У млекопитающих факультативный гетерохроматин проявляется в феномене инактивации Х-хро-
Гилберт с. Биология развития: в 3-х т. Т. 2: Пер. С англ. – м.: Мир, 1994. – 235 с.
ИЗМЕНЕНИЕ ТРАНСКРИПЦИИ В ХОДЕ РАЗВИТИЯ 111
|
Рис. 11.1. Гетерохроматизация отцовских хромосом y самцов мучнистого чернеца. Самец мучнистого чернеца становится функционально гаплоидным, и в сперматозоиде оказываются только эухроматиновые хромосомы. Эти хромосомы становятся гетерохроматиновыми, если потомок окажется самцом |
|
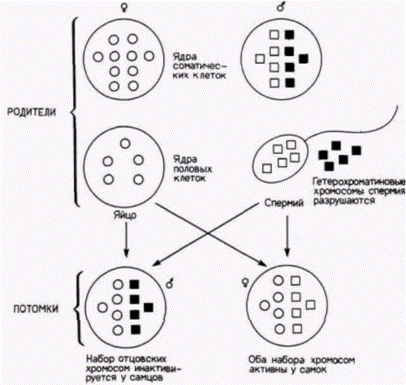
мосомы. В 1949 г. Барр и Бертрам (Barr, Bertram, 1949) обнаружили в ядрах нейронов кошки интенсивно окрашенные тельца, расположенные вблизи ядерной оболочки. Позже эти тельца, названные тельцами Барра, были обнаружены у самок многих млекопитающих, в том числе и у человека: было показано, что они представляют собой неактивную Х-хромосому (рис. 11.2). Это означает, что, хотя в клетках самок содержатся две Х-хромосомы. а в клетках самцов - только одна, у самок транскрипционно активна может быть лишь одна Х-хромосома. Этот феномен называют компенсацией дозы гена. В одной из наиболее ранних работ по инактивации Х-хромосомы изучали закономерности окраски мышей (Магу Lyon, 1961). Если пигментация волосяного покрова определяется аутосомным геном, то окраска мыши будет либо такой, как у одного из родителей, либо промежуточной. В любом случае мышь будет одноцветной. Однако если самка мыши гетерозиготна по Х-сцепленному гену окраски, то результат будет совершенно иной: она будет пятнистой (рис. 11.3). Для объяснения этого феномена исследовательница предложила следующую гипотезу: 1) В ходе раннего развития самок млекопитающих обе Х-хромосомы активны. 2) Впоследствии
|
|
|
Рис. 11.2. Ядра эпителиальных клеток гортани человека, окрашенные фиолетовым крезилом. А. Клетка нормального самца ΧΥ не содержит тельца Барра. Б. Клетка нормальной самки XX содержит одно тельце Барра (указано стрелкой). В. Клетка самки с тремя Х-хромосомами. Можно видеть два тельца Барра: только одна Х-хромосома активна в клетке. (Из Moore, 1977.) |
Гилберт с. Биология развития: в 3-х т. Т. 2: Пер. С англ. – м.: Мир, 1994. – 235 с.
112_______________ ГЛАВА 11______________________________________________________________________________
|
|
Рис 11.3. Инактивация Х-хромосомы у млекопитающих. А. Схема, иллюстрирующая случайную инактивацию Х-хромосомы. Предполагается, что инактивация происходит в период имплантации. Б. Самка мыши, гетерозиготная по гену окраски dappled, связанному с Х-хромосомой. Видны участки с резко различающейся пигментацией. (Фотография с любезного разрешения М. F. Lyon.) |
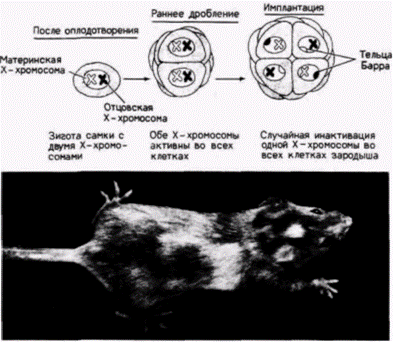
одна из Х-хромосом в каждой клетке выключается.
Эта инактивация происходит случайным образом. В одних клетках инактивируется отцовская Х-хромосома, тогда как в других – материнская.
3) Этот процесс является необратимым. Коль скоро Х-хромосома инактивировалась, она будет неактивной у всех потомков этой клетки. (Участки пигментации у этих мышей достаточно большие.) Таким образом, все ткани у самок млекопитающих являются мозаиками из клеток двух типов.
Некоторые из наиболее убедительных данных в пользу этой модели были получены в биохимических экспериментах с клонами клеток человека. Существует наследственная болезнь человека, называемая синдромом Леша–Найхана; эта болезнь характеризуется отсутствием фермента гипоксантинфосфорибозилтрансферазы (ГФРТ), синтез которого контролируется Х-хромосомой. Синдром Леша–Найхана передается с Х-хромосомой. Это означает, что мужчины, несущие соответствующую мутацию в своей единственной Х-хромосоме, заболевают и умирают от этой болезни. У женщин присутствие мутантного гена ГФРТ может маскироваться другой Х-хромосомой, которая несет аллель дикого типа. Женщина, у которой имеются сыновья, страдающие этой болезнью, является переносчиком, поскольку у нее имеется мутантный ген ГФРТ на одной Х-хромосоме и ген ГФРТ дикого типа на другой Х-хромосоме. Если представленная выше гипотеза правильна, то у такой женщины клетка должна синтезировать либо активный, либо неактивный фермент ГФРТ в зависимости от того, какая Х-хромосома функционирует. Для проверки этого положения индивидуальные клетки кожи, взятые от женщины, гетерозиготной по гену ГФРТ, помещали в культуральную жидкость (Barbara Migeon, 1971). Каждая из этих клеток делилась, образуя клон. При окрашивании этих клонов на присутствие ГФРТ дикого типа было выявлено, что примерно половина клонов содержала этот фрагмент, а половина - нет (рис. 11.4).
Еще один ген Х-хромосомы кодирует глюкозо-6-фосфатдегидрогеназу (Г6ФДГ). У этого фермента имеются обычно два электрофоретических варианта: Г6ФДГ-А и Г6ФДГ-В. Для мужчин характерен вариант А либо В, тогда как женщины в отношении фенотипа по этому ферменту могут быть А, В или AB. В клетках кожи гетерозиготных женщин обнаруживаются оба варианта Г6ФДГ (рис. 11.5). Однако изолированные клоны, полученные после клонирования индивидуальных клеток кожи от этих гетерозигот, экспрессируют только один из двух возможных вариантов. Ни один из клонов не экспрессировал оба варианта (Davidson et al., 1963).
Гипотеза об инактивации Х-хромосомы прекрасно объясняет дифференциальную инактивацию генов на уровне транскрипции. О ее важности дополнительно свидетельствовали некоторые интересные исключения из общих правил. Во-первых, гипотеза оказалась справедливой в отношении сома-