

Гилберт с. Биология развития: в 3-х т. Т. 2: Пер. С англ. – м.: Мир, 1994. – 235 с.
16________________ Глава 7______________________________________________________________________________
|
|
Рис. 7.9. Локализация мРНК в вегетативной шапочке ооцита Xenopus. Ооциты на разных стадиях развития инкубировали с радиоактивной РНК, комплементарной Br1 мРНК. Присутствие и локализацию Brl можно было обнаружить по наличию радиоактивности в любой области яйца (поскольку радиоактивное излучение выявляется с помощью фотографической эмульсии). В ранних ооцитах {Α) Br1 мРНК распределена равномерно по всей цитоплазме. Однако по мере созревания ооцита (Б Г) Brl мРНК постепенно локализуется в области вегетативного полюса яйца Темная область вблизи центра ооцитов зародышевый пузырек. (Из Melton. 1987: фотографии с любезного разрешения D. Melton.) |


![]()
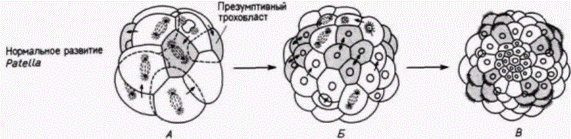
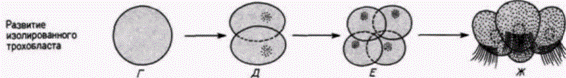
В начале века выдающийся американский эмбриолог Вильсон опубликовал результаты интереснейших опытов. Он изолировал ранние бластомеры зародышей моллюска Patella coerulea и сравнивал их развитие с развитием таких же клеток, оставленных в других зародышах. На рис. 7.10 приведены некоторые результаты опытов, опубликованные Вильсоном в 1904 г. Изолированные бластомеры не только дифференцировались соответственно их проспективному значению (в этом случае формировались реснитчатые клетки трохобласта). но и осуществляли то же самое число клеточных делений и точно в то же самое время, как это делали такие же клетки, оставшиеся в интактном контрольном зародыше. Ориентация этих делений дробления была правильной, и в соответствующее время образовавшиеся клетки формировали реснички. На основании результатов своих опытов Вильсон пришел к выводу, что сами клетки содержат факторы, детерминирующие форму и ритм делений дробления, а сложная дифференцировка. которую они претерпевают, не зависит полностью от их связей с остальной частью зародыша.
|
|
|
Рис. 7.10. Дифференцировка клеток трохобласта у нормального зародыша моллюска Patella (А В) и изолированных клеток трохобласта. культивируемых in vitro (Г Ж). А. Стадия 16 клеток, вид сбоку; презумптивные клетки трохобласта выделены серым цветом. Б. Стадия 48 клеток. В. Личинка, несущая реснички (яиц с анимального полюса). Реснички видны в клетках трохобласта. Г. Изолированная клетка трохобласта. Д. Е. Результаты первого и второго делений в культуре клеток. Ж. Образование ресничек на поверхности изолированных клеток, показанных на рис. 7.10, Е; даже в культуре изолированных клеток появление ресничек по времени точно соответствует их появлению у контрольных зародышей. (По Wilson, 1904.) |
Гилберт с. Биология развития: в 3-х т. Т. 2: Пер. С англ. – м.: Мир, 1994. – 235 с.
__________________ ДЕТЕРМИНАЦИЯ ПОСРЕДСТВОМ ЦИТОПЛАЗМАТИЧЕСКОЙ СПЕЦИФИКАЦИИ____________ 17
Полярная лопасть
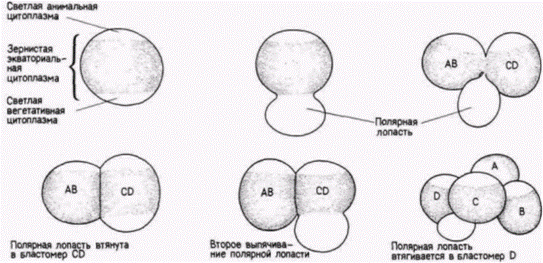
В своем следующем опыте Вильсон сумел покатать, что такое развитие предопределено сегрегацией (распределением) специфических морфогенетических детерминантов по специфическим бластомерам. У зародышей некоторых животных со спиральным дроблением (главным образом у моллюсков и кольчатых червей) непосредственно перед первым делением дробления образуется вырост цитоплазмы (рис. 7.11). Этот вырост называют полярной лопастью. У некоторых видов улиток участок, соединяющий полярную лопасть с яйцом, представляет собой тонкую перемычку. Первое деление дробления делит зиготу асимметрично таким образом, что полярная лопасть соединяется только с бластомером CD. У некоторых видов почти одна треть всего объема цитоплазмы сосредоточена в этой безъядерной лопасти, что придает ей вид отдельной клетки. Стадию двух бластомеров. на которой зародыш имеет такую трехдольную структуру, часто называют трилистником (рис. 7.12). Затем полярная лопасть втягивается в бластомер CD, но снова образуется перед вторым делением (рис. 7 11). После этого деления полярная лопасть оказывается прикрепленной только к бластомеру D, в который она и втягивается. В дальнейшем полярная лопасть более не образуется.
Вильсон показал, что если удалить полярную лопасть на стадии трилистника, оставшиеся клетки делятся нормально. Однако вместо образования нормальной трохофоры (личинки улитки) развивается неполная личинка, совершенно лишенная мезодермальных органов мышц, рта, раковинной железы1 и ноги. Кроме того, Вильсон обнаружил, что тот же самый тип аномального зародыша может быть получен в результате удаления бластомера D на 4-клеточной стадии. На основе этого он пришел к выводу, что цитоплазма полярной лопасти содержит детерминанты мезодермы, которые и придают бластомеру D его способность формировать мезодерму. Вильсон также показал, что локализация мезодермальных детерминантов устанавливается вскоре после оплодотворения и что специфическая цитоплазматическая область яйца, предназначенная быть включенной в бластомер D, содержит какие-то «факторы», необходимые для установления особого ритма дробления бластомера D и для дифференцировки мезодермы.
Морфогенетические детерминанты, содержащиеся в полярной лопасти, по-видимому, локализованы в цитоскелете или в кортикальном слое, а не в жидкой части цитоплазмы зародыша. Клемент (Clement, 1968) центрифугировал зародышей улитки Ilyanassa obsoleta на стадии трилистника, вызывая перетекание жидкой части цитоплазмы обратно в бластомер CD. Однако, когда он затем отрезал полярную лопасть, у сформировавшегося зародыша опять отсутствовали производные мезодермы. Ван ден Биггелаар получил сходные результаты, удаляя цитоплазму из полярной лопасти микропипеткой. Жидкая цитоплазма из других областей клетки
|
|
|
Рис. 7.11. Дробление у моллюска Dentalium. Образование полярной лопасти и ее втягивание происходят дважды. (Из Wilson. 1904.) |
![]()
1 Раковинная железа эктодермальный орган, формирующийся благодаря индукционному влиянию мезодермальных клеток. Без мезодермы в зародыше нет клеток, способных индуцировать компетентную эктодерму. Таким образом, мы снова встречаемся с ограниченной индукцией у мозаичного зародыша.