

Гилберт с. Биология развития: в 3-х т. Т. 2: Пер. С англ. – м.: Мир, 1994. – 235 с.
__________________ ТРАНСЛЯЦИОННАЯ И ПОСТТРАНСЛЯЦИОННАЯ РЕГУЛЯЦИЯ РАЗВИТИЯ____________________ 213
|
|
|
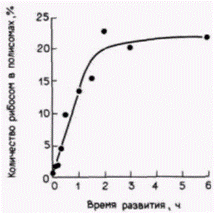
Рис. 14.18. Увеличение относительного количества рибосом, включенных в состав полисом, в ходе раннего развития морского ежа. (По Humphreys, 1971.) |
шиеся фракции мРНК. Молекулы мРНК для трех известных белков, специфичных для зародышей (А, В и С), в цитоплазме ооцита обнаруживаются в составе нетранслируемых РНК, тогда как в бластомерах зародыша они обнаруживаются в полисомной мРНК Напротив, молекулы мРНК для трех белков, характерных для ооцитов (Χ, Y и Z), выявляются в полисомах ооцитов, но не зародышей. Таким образом, можно проследить рекрутирование мРНК из нетранслируемых РНП в транскрипционно активные полисомы.
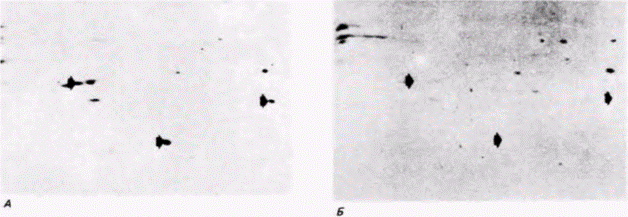
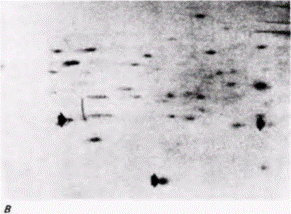
Рекрутирование мРНК наблюдается также у зародышей Drosophila. Из РНП и полисом ооцита, а также полисом зародышей дрозофилы выделили поли(А)-содержащую мРНК, которую затем транслировали в бесклеточной системе, содержащей радиоактивные аминокислоты (Mermod et al., 1980). Было обнаружено, что на мРНК из РНП ооцитов можно синтезировать определенные белки, которые нельзя синтезировать на мРНК из полисом ооцитов (рис. 14.19, А, Б). Однако эти белки можно синтезировать на мРНК, выделенной из полисом зародыша (рис. 14.19.0). Следовательно, мРНК, первоначально запасенная в РНП-частицах ооцита, вовлекается в полисомы зародыша. Таким способом инициация трансляции контролирует экспрессию генов.
мРНК БЕЗ КЭП-ГРУППЫ. Для эффективной трансляции необходимы 5'- и 3'-концы мРНК. Мы уже видели, как различия в длине 3’-поли(А)-хвостов
|
| |
|
|
Рис. 14.19. Данные, свидетельствующие о вовлечении РНК из РНП ооцита в полисомы зародыша. Поли(А)-содержащую РНК выделили из РНП ооцитов (А), полисом ооцитов (Б) и полисом зародышей (В). Полученную РНК транслировали in vitro в присутствии радиоактивных аминокислот и затем реакционную смесь пропускали через колонку с ДНК для получения набора белков, способных связаться с ДНК. Эти белки элюировали с колонки и подвергали изофокусированию в первом направлении (для разделения белков по электрическому заряду) и затем электрофорезу во втором направлении (для разделения белков по массе). На радиоавтографах двумерных гелей видны три белка, которые синтезируются на РНК ооцитных РНП и позже на мРНК полисом зародышей (указаны стрелками на А и В). Эти три белка не синтезируются на РНК, экстрагированных из полисом ооцитов (указаны стрелками на Б). (Из Mermod et al.,. 1980; с любезного разрешения J. Mermod.) |
Гилберт с. Биология развития: в 3-х т. Т. 2: Пер. С англ. – м.: Мир, 1994. – 235 с.
214_______________ ГЛАВА 14____________________________________________________________________________
|
| |
|
|
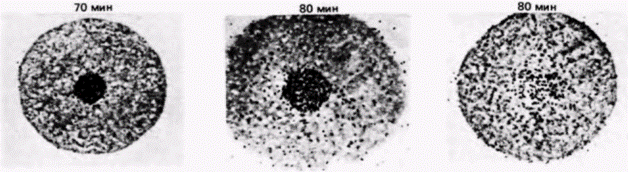
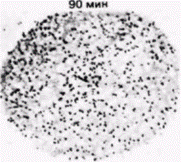
Рис. 14.20. Компартментализация мРНК гистонов в ооците морского ежа. Зонд кДНК, узнающий гистоновую мРНК. добавляли к яйцеклеткам морского ежа, фиксированным в различные сроки после оплодотворения. Радиоавтографы показывают, что данная мРНК локализована в материнском пронуклеусе вплоть до его дезинтеграции через 80–90 мин после проникновения спермия. (Из De Leon et al., 1983; фотографии с любезного разрешения L. и R. Angerer.) |
могут влиять на дифференциальную трансляцию РНК в ооцитах Spisula. Некоторые бабочки используют для трансляционного контроля механизм, связанный с изменениями 5'-кэп-группы (Kastern et al., 1982). Чтобы эффективно транслироваться, почти все эукариотические мРНК нуждаются в присутствии на своих 5'-концах кэп-группы, содержащей 7-метилгуанозин (Shatkin, 1976). В ооците табачного бражника запасенные мРНК имеют неметилированную кэп-группу, при этом гуанозин содержится, но к нему не добавлена метильная группа. Такие мРНК не транслируются в белки в бесклеточной системе. Однако при оплодотворении в ооцитах бражника проходит волна метилирования и завершается формирование кэп-групп. Молекулы мРНК, обладающие сформированными кэп-группами, способны затем связаться с рибосомами и инициировать трансляцию. В любом случае, замаскирована ли молекула мРНК белками или не завершена модификация ее 5’-конца, инициация трансляции будет подавлена. Эта молекула может не транслироваться до тех пор. пока не поступит соответствующий сигнал.
КОМПАРТМЕНТАЛИЗОВАННЫЕ мРНК. В некоторых работах подвергается сомнению вывод о том, что ооцитные РНП не способны к трансляции. Вместо этого полагают, что белоксинтезирующий аппарат в ооците компартментализован, благодаря чему закрыт доступ мРНК (в составе РНП) к рибосомам (Moon et al., 1982). Синтез гистоновых мРНК в ооцитах морского ежа регулируется, по-видимому, с помощью разграничения такого типа. Гистоновые мРНК локализованы не в цитоплазме ооцита, а в его большом пронуклеусе. И только по завершении оплодотворения, когда пронуклеус разрушается, гистоновая мРНК выходит в цитоплазму (рис. 14.20) (DeLeon et al., 1983). Этого может не быть в случае других мРНК. В пронуклеусе находится менее 0,1% всей мРНК ооцита (Angerer, Angerer, 1981), а те РНП, которые содержат мРНК для актина и тубулина, локализованы преимущественно в цитоплазме (Showman et al., 1982). Известно, что некоторые материнские мРНК, как и отдельные рибосомы, присоединены к цитоскелету (Moon et al., 1983), следовательно, цитоскелет также может отделить молекулы мРНК от рибосом.
ИЗМЕНЕНИЕ ЭФФЕКТИВНОСТИ ТРАНСЛЯЦИИ. В предложенных ранее моделях трансляционной регуляции предполагается, что транскрипционный аппарат способен эффективно транслировать любую мРНК, но что мРНК и рибосомы физически или химически отделены друг от друга. На самом деле этого не требуется. Исходно низкое значение pH в ооците само по себе может препятствовать синтезу белка. Как обсуждалось в гл. 2, у морского ежа в ходе оплодотворения происходит значительное высвобождение ионов водорода, приводящее к увеличению значения pH в цитоплазме с 6,9 до 7,4. Из неоплодотворенных ооцитов морского ежа группой исследователей была получена бесклеточная систе-