

Гилберт с. Биология развития: в 3-х т. Т. 2: Пер. С англ. – м.: Мир, 1994. – 235 с.
|
|
|
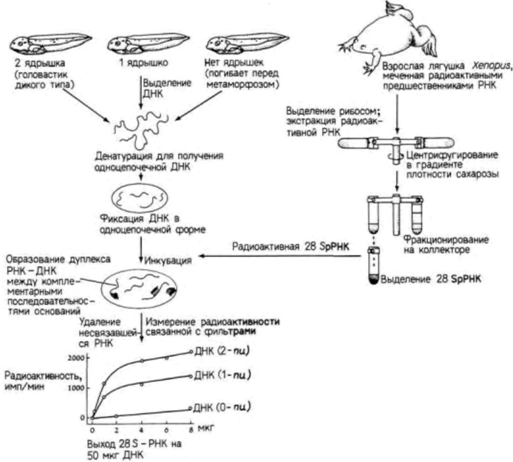
Рис. 10.1. Опыт по гибридизации нуклеиновых кислот, из которого становится ясно, что ядрышко кодирует рибосомную 28S-PHK. Из содержащих 2, 1 и 0 ядрышек клеток головастиков экстрагировали по отдельности ДНК. Полученную ДНК денатурировали (делали одноцепочечной) и определенное количество одноцепочечной ДНК запекали на нитроцеллюлозных фильтрах. Одновременно из клеток взрослых лягушек получали радиоактивную 28S-pPHK путем выделения рибосом и последующего разделения трех основных видов РНК. Радиоактивную 28S-pPHK инкубировали на ДНК-содержащих фильтрах в условиях, когда она могла присоединяться к комплементарным участкам ДНК. Затем измеряли радиоактивность фильтров в сцинтилляционном счетчике. |
|
|
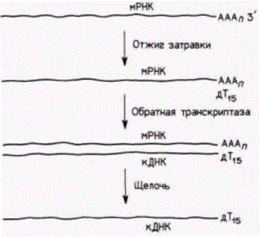
Рис. 10.2. Метод получения комплементарной ДНК (кДНК). Большая часть информационных РНК (мРНК) содержит протяженную последовательность из остатков аденозина (АААn) на 3’-концс молекул (см. гл. 11): поэтому, экспериментаторы гибридизуют «затравку», состоящую из 15 остатков дезокситимидина (дТ15), с 3'-концом мРНК. Затем с помощью обратной транскриптазы синтезируют комплементарную цепь ДНК, начиная от затравки дТ15., кДНК можно выделить с помощью повышения pH раствора, что вызывает денатурацию двухцепочечного гибрида и деградацию РНК. |
Гилберт с. Биология развития: в 3-х т. Т. 2: Пер. С англ. – м.: Мир, 1994. – 235 с.
________ ТОЖДЕСТВО ГЕНОМОВ И ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ 83
|
Таблица 10.1. Широко используемые рестриктазы | ||
|
Фермент |
Источник |
Сайт узнавания и расщепления1 |
|
| ||

1 Все сайты узнавания рестриктаз имеют центральную симметрию. Двухцепочечная последовательность, прочитанная в одном направлении, идентична последовательности, прочитанной в обратном направлении.
На рис. 10.3 показана плазмида pBR322, которая часто используется молекулярными биологами в качестве клонирующего вектора. Она несет два гена устойчивости к антибиотикам – tcR, который обеспечивает устойчивость к тетрациклину, и АрR, который делает бактерии устойчивыми к ампициллину. Эти гены устойчивости к антибиотикам, столь нежелательные для медиков, становятся незаменимыми для генных инженеров, поскольку один из BamH1 сайтов в этой плазмиде расположен в середине гена tcR. Предполагаемые рекомбинантные плазмиды. полученные описанным ранее способом, затем инкубируют с клетками Ε. coli дикого типа, чувствительными к обоим антибиотикам, в условиях, позволяющих бактериям захватить эти плазмиды. При этом не каждая бактерия включает плазмиду и не каждая плазмида включает чужеродный ген. т.к. липкие концы плазмиды в сайте рестрикции могут ренатурировать друг с другом. Для отбора тех бактерий, которые включили плазмиды, обработанные клетки Ε. coli выращивают на агаре в присутствии ампициллина. При этом выживают только те бактерии, которые включили плазмиды. Затем часть каждой жизнеспособной колонии переносится на агар, содержащий тетрациклин. Если бактерии продолжают рост, то, следовательно, ген tcR плазмиды остался интактным и эта плазмида восстановилась в результате простого соединения собственных концов друг с другом. Если бактерии, выросшие на среде с ампициллином, не выживают на среде с тетрациклином, то ген устойчивости к тетрациклину в плазмиде уже не функционирует. Это означает, что ген tcR разорван фрагментом фермента рестрикции из чужого генома. Со среды, содержащей ампициллин, отбираются те колонии, которые не росли на среде, содержащей тетрациклин. Эти колонии следует проверить на присутствие определенного гена.
Клетки из каждой колонии E. coli, содержащей рекомбинантные плазмиды, помещают на нитроцеллюлозный фильтр. После лизиса клеток клеточная ДНК присоединяется к фильтру. Затем цепи ДНК разделяют с помощью нагревания и инкубируют фильтр в растворе, содержащем радиоактивную РНК (или ее кДНК-копию) для гена, выбранного для клонирования. Если плазмида содержит этот ген, то ДНК этого гена должна присутствовать на бумаге и только эта ДНК обладает способностью присоединять радиоактивную РНК или кДНК. Именно поэтому соответствующие места фильтра окажутся радиоактивными. Радиоактивность этих участков регистрируется с помощью радиоавтографии. Для этого чувствительная рентгеновская пленка помещается поверх обработанной бумаги. Электроны с высокой энергией, излучаемые радиоактивной РНК, сенсибилизируют зерна серебра в пленке, вызывая их потемнение при последующем проявлении. В результате над каждой колонией, содержащей рекомбинантную плазмиду с определенным геном, образуется темное пятно (рис. 10.3). Эти колонии изолируют и выращивают вновь. В итоге можно получить многие триллионы клеток, каждая из которых содержит сотни рекомбинантных плазмид.
Рекомбинантные плазмиды можно отделить от хромосомы Ε. coli с помощью центрифугирования. Фрагмент ДНК человека, содержащий интересующий нас ген, высвобождается при инкубации плазмид с BamH1. Этот фрагмент затем отделяют от плазмидной ДНК. Таким образом, экспериментатор получает микрограммы очищенной ДНК, содержащей определенный ген. [Хотя описанная процедура выглядит вполне логичной и простой, число колоний, которые следует скринировать, достигает часто астрономических чисел. Число случайных фрагментов, которые должны быть клонированы, для того чтобы получить интересующий вас ген, возрастает с увеличением сложности генома организма1. Чтобы клонировать с 99%-ной надежностью желаемый ген из Е. coli, необходимо проанализировать примерно 1500 колоний. Для млекопитающих это число возрастет до 800 000: вместо
![]()
1 Сложность в данном случае определяется числом генов различного типа, содержащихся в клеточном ядре.