

Сплайсинг белков
Феномен сплайсинга белков, обнаруженный в 1990 г. в группой Т.Стивенса, пошатнул еще один постулат молекулярной биологии, в соответствии с которым последовательности нуклеотидов зрелых мРНК всегда колинеарны аминокислотным последовательностям кодируемых ими полипептидов. Во время сплайсинга белков удаление избыточной генетической информации из макромолекул происходит не на уровне пре-мРНК, как это имеет место во время обычного сплайсинга, а на уровне синтезированного полипептида путем вырезания из его внутренней части короткой аминокислотной последовательности. Именно данный механизм отличает сплайсинг белков от повсеместно распространенного процессинга предшественников полипептидов, который, как известно, сопровождается только протеолитическим расщеплением полипептида-предшественника с образованием более коротких белков без изменения их внутренней первичной структуры.
Открытие сплайсинга белков было сделано при исследовании экспрессии гена VMA1 дрожжей S. cerevisiae, известного также под названием TFP1, который кодирует субъединицу ATPазы вакуолей с молекулярной массой 119 кДа (рис. I.43,а). При исследовании гомологии данного гена с генами других ATPаз микроорганизмов было установлено, что все они кодируют более короткие полипептиды с молекулярными массами 70 кДа, и их гомология с геном дрожжей распространяется только на концевые последовательности нуклеотидов, резко нарушаясь в центральной части гена. Более того, использование инсерционного мутагенеза (генного нокаута) (см. раздел 9.1.3) для инактивации этого гена приводило к прекращению синтеза в клетках дрожжей полипептида с молекулярной массой 69, а не 119 кДа. Механизм такого явления прояснился после постановки двух контрольных экспериментов. В первом из них введение мутаций со сдвигом рамки считывания в сегмент гена VMA1, который кодировал центральную часть полипептида, отсутствующую в зрелом белке, приводило к прекращению синтеза всего белка рибосомами из-за возникновения новых терминирующих кодонов в этой рамке считывания мРНК. Подобное бы не происходило, если бы центральная часть пре-мРНК гена VMA1 удалялась в результате сплайсинга. Во втором эксперименте исследование экстрактов клеток дрожжей с помощью антител к центральной части полипептида, отсутствующего в зрелой субъединице, обнаружило белковый продукт с предсказанной молекулярной массой 50 кДа. Он присутствовал в соотношении 1:1 к функционально активному полипептиду с молекулярной массой 69 кДа. Это указывало на то, что последовательности РНК центральной части гена транслируются рибосомами и соответствующая часть полипептида-предшественника удаляется посттрансляционно. Дальнейшие исследования данного явления полностью подтвердили предположение о том, что внутренняя аминокислотная последовательность из предшественника с молекулярной массой 119 кДа отщепляется в результате сплайсинга на уровне его полипептидной цепи (см. рис. I.43,а). Внутренняя часть полипептида, удаляемая в результате белкового сплайсинга, получила название интеина, а наружные N- и C-концевые части – экстеинов. Позднее белки, изменяемые посттрансляционно в результате белкового сплайсинга, были обнаружены у многих микроорганизмов (см. рис. I.43,б). Во всех случаях аминокислотные последовательности интеинов фланкированы короткими консервативными последовательностями. На основании особенностей первичной структуры интеинов и экстеинов разработаны алгоритмы поиска белков, подвергаемых сплайсингу посттрансляционно, в соответствующих базах данных. С использованием этого алгоритма, в частности обнаружена последовательность интеина в продукте гена dnaB хлоропластов красной водоросли Porphira porphira.
Для объяснения механизма белкового сплайсинга предложено несколько моделей. Одна из них, разработанная группой Ф. Перлера и включившая в себя многие черты ранних моделей, представлена на рис. I.44. В соответствии с этой моделью белковый сплайсинг является аутокаталитическим процессом и для своего осуществления не требует других кофакторов в дополнение к самому полипептиду-предшественнику. Событием, запускающим белковый сплайсинг, может быть: а) нуклеофильная атака OH-группой консервативного остатка Ser, расположенного в C-концевом сайте сплайсинга, по карбонильной группе пептидной связи, расположенной в N-концевом сайте сплайсинга, или же: б) N–O-сдвиг в N-концевом сайте сплайсинга с последующей атакой OH-группой C-концевого сайта и переэтерификацией. В результате функционирования обоих механизмов образуется разветвленное промежуточное соединение, которое при участии остатка Asn распадается на интеин-сукцинимид и экстеины, связанные друг с другом сложноэфирной связью. Эта связь преобразуется в пептидную в результате N–O-сдвига.
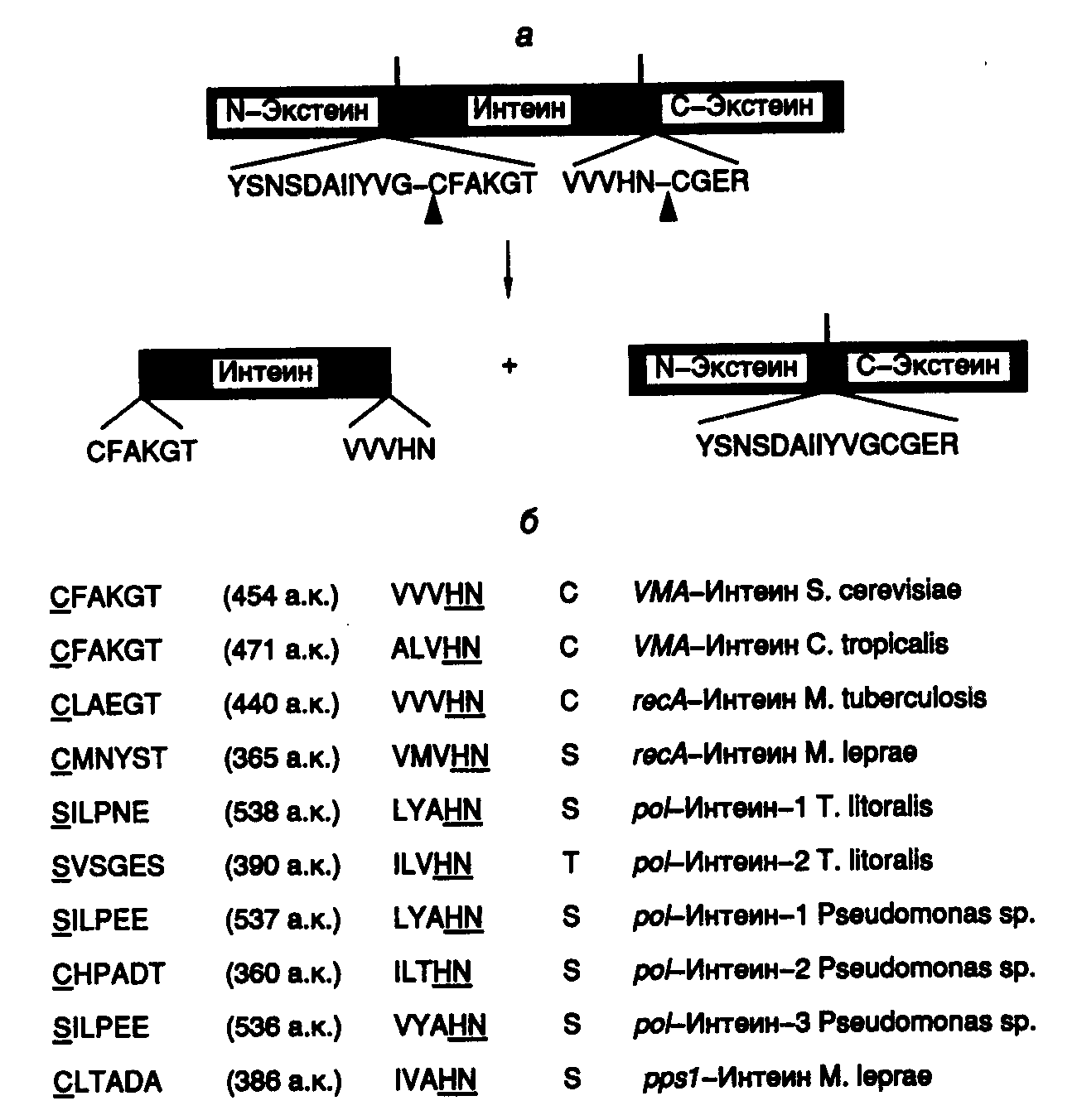
Рис. I.43. Сплайсинг белка гена TFP1 S. cerevisiae (а) и перечень генов, белки которых подвергаются сплайсингу на уровне полипептидных цепей (б)
Стрелки указывают места разрыва полипептидной цепи белка-предшественника TFP1. Консервативные последовательности 10 известных интеинов в окрестностях сайтов сплайсинга изображены с использованием однобуквенного кода
Генетическая мобильность последовательностей интеинов в ДНК является одной из самых больших неожиданностей, обнаруженных после их открытия. Оказалось, что полипептидная цепь интеина обладает эндонуклеазной активностью, которая обеспечивает транспозиции последовательностей интеинов в геноме. Аналогичный процесс, ранее получивший название "хоуминга интронов", описан у интронов группы I. В результате хоуминга происходит однонаправленный перенос копии последовательности ДНК интрона (или интеина) из гена, содержащего эту последовательность, к аллельному гену, не содержащему ее. Данная цепь реакций инициируется эндонуклеазным разрывом обеих цепей ДНК в аллельном безинтеиновом гене, образующемся под действием эндонуклеазы интеина. Далее с использованием последовательности ДНК интеина в качестве донора информации происходит репарация двухцепочечного разрыва, которая сопровождается конверсией гена с включением в его состав последовательности интеина. Это указывает на большое сходство механизмов хоуминга интронов группы I и интеинов.
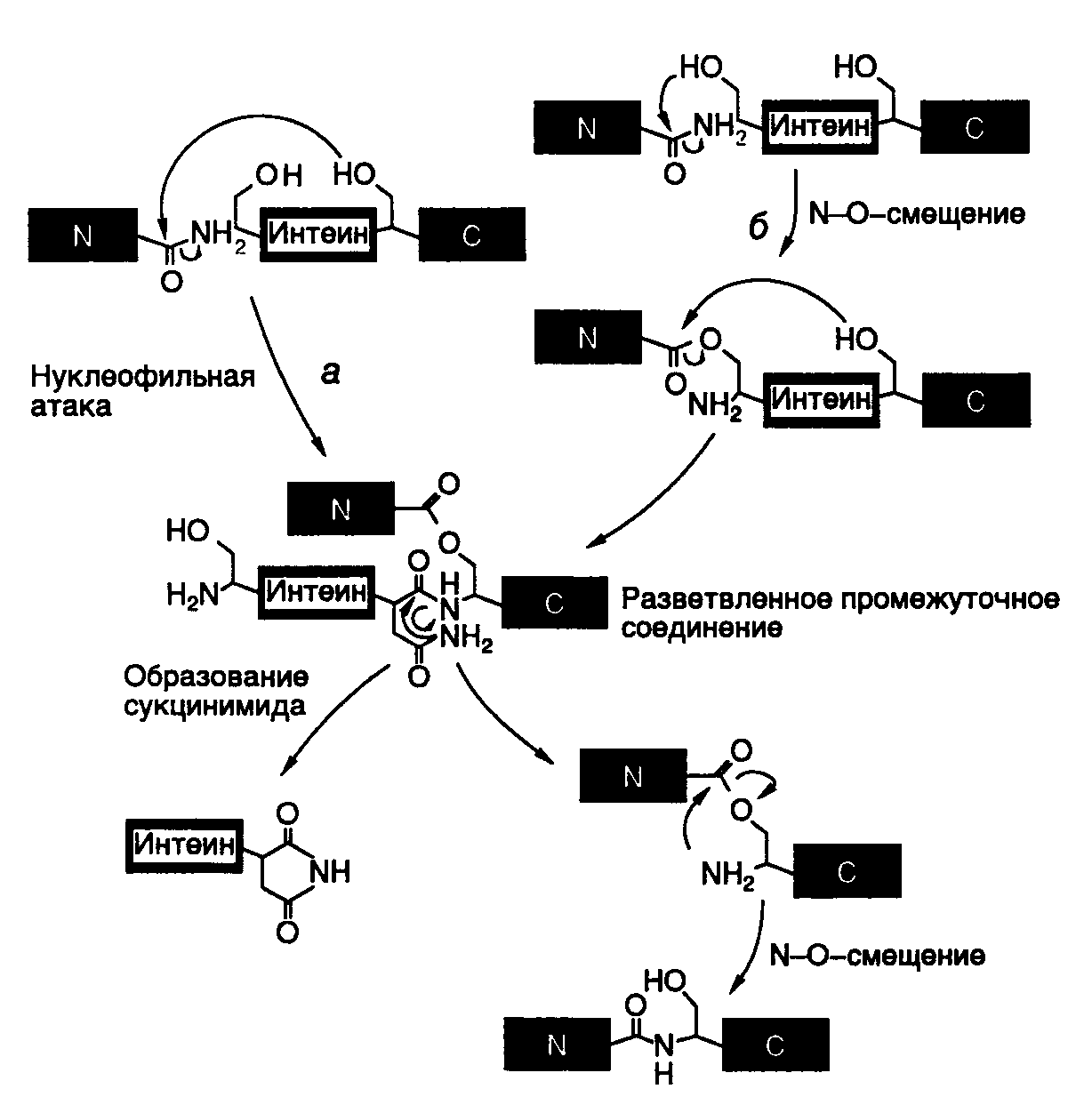
Рис. I.44. Предполагаемый механизм сплайсинга белков
а, б– альтернативные механизмы инициации сплайсинга, N, C – N- и C-концевые экстеины белков-предшественников
Сплайсинг белков является сложной задачей для исследования in vitro. Процесс вырезания последовательности интеина происходит настолько быстро, что не удается обнаружить промежуточные соединения. Проблему удалось решить методами генной инженерии, вставив последовательность ДНК интеина из гена ДНК-полимеразы термофильной бактерии Pyrococcus в одну рамку считывания между геном белка, связывающего мальтозу, и частью гена парамиозина Dirofilaria immitis. Образующийся химерный белок не претерпевал сплайсинга при 12–20°, но при 37–65° сплайсинг индуцировался. Именно с помощью этого подхода удалось обнаружить разветвленные промежуточные соединения, образующиеся при сплайсинге белков. Кроме того, такой подход может стать продуктивным для биотехнологии, так как он позволяет нарабатывать токсические для клетки белки в виде химерных предшественников при низких температурах, которые затем простым повышением температуры быстро превращаются в полезные белковые продукты.