

Хроматин
Хроматином называют сложную смесь веществ, из которых построены хромосомы эукариот. Основными компонентами хроматина являются ДНК, гистоны и негистоновые белки, образующие высокоупорядоченные в пространстве структуры. Соотношение ДНК и белка в хроматине составляет ~1:1, а основная масса белка хроматина представлена гистонами. Гистоны образуют семейство высококонсервативных основных белков, которые разделяются на пять больших классов, названных H1, H2A, H2B, H3 и H4. Размер полипептидных цепей гистонов лежит в пределах ~220 (H1) и 102 (H4) аминокислотных остатков. Гистон H1 сильно обогащен остатками Lys, для гистонов H2A и H2B характерно умеренное содержание Lys, полипептидные цепи гистонов H3 и H4 богаты Arg. Внутри каждого класса гистонов (за исключением H4) на основании аминокислотных последовательностей различают несколько субтипов этих белков. Такая множественность особенно характерна для гистонов класса H1 млекопитающих. В этом случае различают семь субтипов, названных H1.1–H1.5, H1o и H1t.
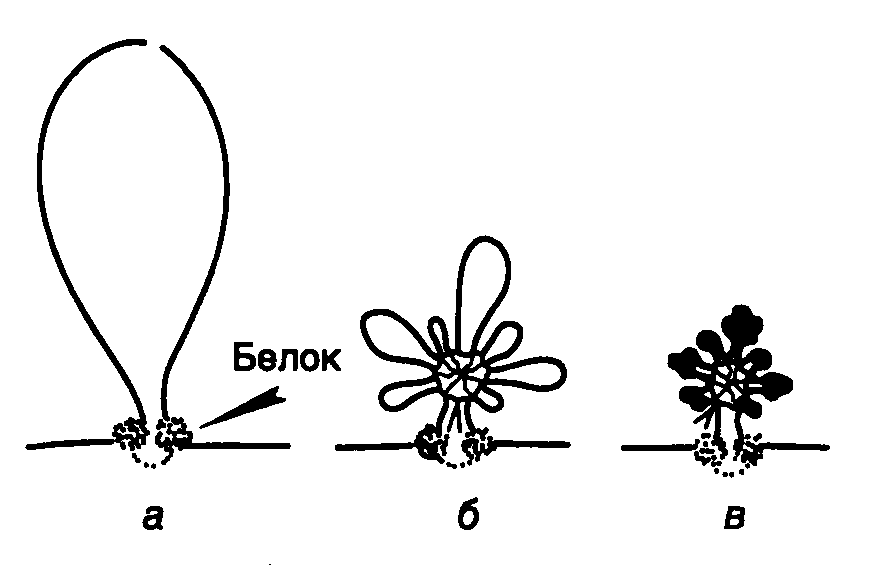
Рис. I.2. Схематическое изображение петельно-доменного уровня компактизации хроматина
а– фиксация петли хромомера на ядерном матриксе с помощью MAR/SAR-последовательностей и белков;б– "розетки", образованные из петли хромомера;в– конденсация петель "розеток" с участием нуклеосом и нуклеомеров
Важным результатом взаимодействия ДНК с белками в составе хроматина является ее компактизация. Суммарная длина ДНК, заключенной в ядре клеток человека, приближается к 1 м, тогда как средний диаметр ядра составляет 10 мкм. Длина молекулы ДНК, заключенной в одной хромосоме человека, в среднем равняется ~4 см. В то же время длина метафазной хромосомы составляет ~4 мкм. Следовательно, ДНК метафазных хромосом человека компактизована по длине, по крайней мере, в 104 раз. Степень компактизации ДНК в интерфазных ядрах значительно ниже и неравномерна в отдельных генетических локусах. С функциональной точки зрения различают эухроматин и гетерохроматин. Эухроматин характеризуется меньшей по сравнению с гетерохроматином компактизацией ДНК, и в нем главным образом локализуются активно экспрессирующиеся гены. В настоящее время широко распространено мнение о генетической инертности гетерохроматина. Поскольку его истинные функции сегодня нельзя считать установленными, эта точка зрения по мере накопления знаний о гетерохроматине может измениться. Уже сейчас в нем находят активно экспрессирующиеся гены.
Гетерохроматизация определенных участков хромосом часто сопровождается подавлением транскрипции имеющихся в них генов. В процесс гетерохроматизации могут быть вовлечены протяженные участки хромосом и даже целые хромосомы. В соответствии с этим считается, что регуляция транскрипции генов эукариот в основном происходит на двух уровнях. На первом из них компактизация или декомпактизация ДНК в хроматине может приводить к длительной инактивации или активации протяженных участков хромосом или даже целых хромосом в онтогенезе организма. Более тонкая регуляция транскрипции активированных участков хромосом достигается на втором уровне при участии негистоновых белков, включающих многочисленные факторы транскрипции.
Структурная организация хроматина и хромосом эукариот. Вопрос о структурной организации хроматина в интерфазных ядрах в настоящее время далек от своего разрешения. Это связано, прежде всего, со сложностью и динамичностью его структуры, которая легко меняется даже при незначительных экзогенных воздействиях. Большинство знаний о структуре хроматина было получено in vitro на препаратах фрагментированного хроматина, структура которого значительно отличается от таковой в нативных ядрах. В соответствии с распространенной точкой зрения различают три уровня структурной организации хроматина у эукариот: 1) нуклеосомная фибрилла; 2) соленоид, или нуклеомер; 3) петельно-доменная структура, включающая хромомеры.
Нуклеосомные фибриллы. В определенных условиях (при низкой ионной силе и в присутствии двухвалентных ионов металлов) в изолированном хроматине удается наблюдать регулярные структуры в виде протяженных фибрилл диаметром 10 нм, состоящих из нуклеосом. Эти фибриллярные структуры, в которых нуклеосомы расположены как бусы на нитке, рассматриваются в качестве низшего уровня упаковки ДНК эукариот в хроматине. Нуклеосомы, входящие в состав фибрилл, расположены более или менее равномерно вдоль молекулы ДНК на расстоянии 10–20 нм друг от друга. В состав нуклеосом входят четыре пары молекул гистонов: H2a, H2b, H3 и H4, а также одна молекула гистона H1. Данные по структуре нуклеосом в основном получены с использованием трех методов: рентгеноструктурного анализа низкого и высокого разрешения кристаллов нуклеосом, межмолекулярных сшивок белок–ДНК и расщепления ДНК в составе нуклеосом с помощью нуклеаз или радикалов гидроксила. На основании таких данных А. Клугом была построена модель нуклеосомы, в соответствии с которой ДНК (146 п.о.) в B-форме (правозакрученная спираль с шагом 10 п.о.) намотана на гистоновый октамер, в центральной части которого расположены гистоны Н3 и Н4, а на периферии – Н2а и Н2b. Диаметр такого нуклеосомного диска составляет 11 нм, а его толщина – 5,5 нм. Структура, состоящая из гистонового октамера и намотанной на него ДНК, получила название нуклеосомной кóровой частицы. Кóровые частицы отделены друг от друга сегментами линкерной ДНК. Общая длина участка ДНК, включенного в нуклеосому животных, составляет 200 (15) п.о.
Полипептидные цепи гистонов содержат структурные домены нескольких типов. Центральный глобулярный домен и гибкие выступающие N- и С-концевые участки, обогащенные основными аминокислотами, получили название плеч (arm). С-концевые домены полипептидных цепей, участвующие в гистон–гистоновых взаимодействиях внутри кóровой частицы, находятся преимущественно в виде -спирали с протяженным центральным спиральным участком, вдоль которого с двух сторон уложено по одной более короткой спирали. Все известные места обратимых посттрансляционных модификаций гистонов, происходящих на протяжении клеточного цикла или во время дифференцировки клеток, локализованы в гибких основных доменах их полипептидных цепей (табл. I.2). При этом N-концевые плечи гистонов H3 и H4 являются самыми консервативными участками молекул, а гистоны в целом – одними из наиболее эволюционно консервативных белков. С помощью генетических исследований дрожжей S. cerevisiae было установлено, что небольшие делеции и точковые мутации в N-концевых частях генов гистонов сопровождаются глубокими и разнообразными изменениями фенотипа дрожжевых клеток. Это указывает на чрезвычайную важность целостности молекул гистонов в обеспечении правильного функционирования эукариотических генов.
В растворе гистоны Н3 и Н4 могут существовать в виде стабильных тетрамеров (Н3)2(Н4)2, а гистоны Н2А и Н2В – в виде стабильных димеров. Постепенное повышение ионной силы в растворах, содержащих нативный хроматин, приводит к освобождению сначала димеров Н2А/Н2В, а затем тетрамеров Н3/Н4.
Дальнейшее уточнение тонкой структуры нуклеосом в кристаллах было проведено недавно в работе К. Люгера с соавт. (1997 г.) с помощью рентгеноструктурного анализа высокого разрешения. Было установлено, что выпуклая поверхность каждого гистонового гетеродимера в составе октамера огибается сегментами ДНК длиной 27–28 п.о., расположенными по отношению друг к другу под углом 140о, которые разделены линкерными участками длиной в 4 п.о.
В соответствии с современными данными пространственная структура ДНК в составе кóровых частиц несколько отличается от B-формы: двойная спираль ДНК перекручена на 0,25–0,35 п.о./виток двойной спирали, что приводит к образованию шага спирали, равному 10,2 п.о./виток (у В-формы в растворе – 10,5 п.о./виток). Стабильность комплекса гистонов в составе кóровой частицы определяется взаимодействием их глобулярных частей, поэтому удаление гибких плеч в условиях мягкого протеолиза не сопровождается разрушением комплекса. N-концевые плечи гистонов, по-видимому, обеспечивают их взаимодействие со специфическими участками ДНК. Так, N-концевые домены гистона Н3 контактируют с участками ДНК на входе в кóровую частицу и выходе из нее, тогда как соответствующий домен гистона Н4 связывается с внутренней частью ДНК нуклеосомы.
Упомянутые выше исследования структуры нуклеосом высокого разрешения показывают, что центральная часть сегмента ДНК длиной в 121 п.о. в составе нуклеосомы образует дополнительные контакты с гистоном H3. При этом N-концевые части полипептидных цепей гистонов H3 и H2B проходят через каналы, образуемые малыми бороздками соседних супервитков ДНК нуклеосомы, а N-концевая часть гистона H2A контактирует с малой бороздкой внешней части супервитка ДНК. В совокупности данные высокого разрешения показывают, что ДНК в составе коровых частиц нуклеосом огибает гистоновые октамеры неравномерно. Кривизна нарушается в местах взаимодействия ДНК с поверхностью гистонов, и такие изломы наиболее заметны на расстоянии 10–15 и 40 п.о. от центра супервитка ДНК.
Таблица I.2