

Рибозимы и дезоксирибозимы
Регуляция экспрессии генов с помощью антисмысловых РНК характеризуется высокой специфичностью. Это обусловлено большой точностью процесса РНК-РНК-гибридизации, основанной на комплементарном взаимодействии друг с другом протяженных последовательностей нуклеотидов. Однако сами по себе антисмысловые РНК не инактивируют необратимо мРНК-мишени, и для подавления экспрессии соответствующих генов требуются высокие (по крайней мере, эквимолярные по отношению к мРНК) внутриклеточные концентрации антисмысловых РНК. Эффективность действия антисмысловых РНК резко повысилась после того, как в их состав были введены молекулы рибозимов – коротких последовательностей РНК, обладающих эндонуклеазной активностью. Хотя ниже термин "рибозимы" будет использоваться почти исключительно в отношении молекул РНК, обладающих эндонуклеазной активностью, необходимо иметь в виду, что в настоящее время известно множество других ферментативных активностей, ассоциированных с РНК. Поэтому рибозимами в широком смысле называют молекулы РНК, обладающие любой ферментативной активностью. Некоторые из этих активностей будут рассмотрены в разделе 9.3 при обсуждении возможной роли РНК в происхождении жизни.
Типы рибозимов
Эндорибонуклеазная активность РНК была впервые обнаружена Т. Чехом в 1980 г. у интрона группы I предшественника рибосомной РНК Tetrahymena, осуществляющего аутокаталитическую реакцию сплайсинга (аутосплайсинга), в результате которой происходит вырезание из молекулы предшественника рРНК последовательности этого интрона и образование зрелой молекулы рРНК. (Молекулярный механизм реакции аутосплайсинга уже был рассмотрен в первой части книги в разделе 3.3.2.) С тех пор аутокаталитические реакции расщепления были выявлены у многих молекул РНК (рис. II.25). Наиболее перспективными для использования в составе антисмысловых РНК оказались рибозимы, образующие пространственную структуру типа "головки молотка" (hammerhead – HH) (см. рис. II.25,а). Такие РНК были найдены у сателлитных РНК вирусов растений, вироидов, а также среди транскриптов сателлитных ДНК тритонов. У РНК подобного типа была определена минимальная каноническая последовательность, необходимая для появления ферментативной активности, длиной в 27 нуклеотидов, из которых высококонсервативны 13 нуклеотидов, а также последовательность из трех нуклеотидов, в пределах которой происходит расщепление фосфодиэфирной связи (см. рис. II.25,а).
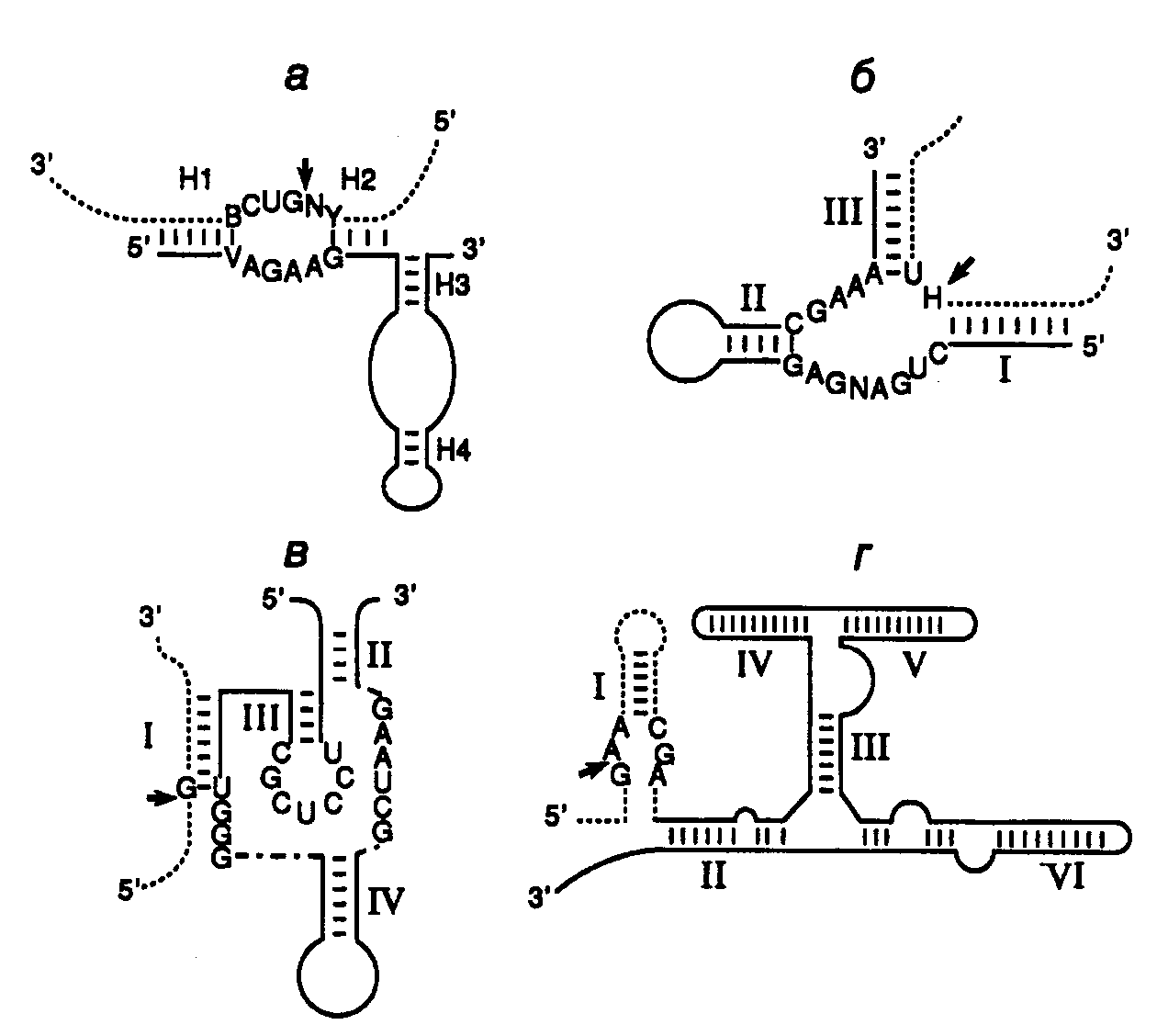
Рис. II.25. Особенности вторичной структуры рибозимов разных типов
а– "головка молотка",б– шпилька,в– рибозим вируса гепатита,г– рибозимNeurosporaVS. Стрелки обозначают точки расщепления РНК; нуклеозид N может быть A, U, G или C, H – A, U или C, Y – любой пиримидин; приведены общепринятые обозначения элементов вторичных структур рибозимов
Другими примерами рибозимов являются РНК, образующие структуры типа шпилек, а также РНК вируса гепатита δ (ВГД) и сателлитная РНК Варкуда (Varcud satellite – VS) нейроспоры (см. рис. II.25,б,в,г соответственно). (Структура и механизм действия рибозимов в составе интронов групп I, II и III были подобно рассмотрены в разделе 2.2.3.) Рибозимы со структурами типа шпилек были впервые обнаружены у вироидов и вирусоидов растений, рибозимы ВГД были идентифицированы среди сателлитных РНК вируса гепатита В человека, а рибозимы VS – среди сателлитных РНК природных изолятов Neurospora. В связи с рибозимами следует вспомнить и о РНКазе Р E. coli, состоящей из небольшого полипептида и РНК длиной в 375 нуклеотидов, о которой уже упоминалось в разделе 2.2.1. Именно РНК обеспечивает этому ферменту эндонуклеазную активность, необходимую для процессинга предшественников тРНК.
Хотя у природных РНК сайт расщепления и последовательность нуклеотидов рибозима расположены на одной молекуле (in cis по отношению друг к другу), HH-рибозимы могут действовать и на сайты расщепления, расположенные in trans, т.е. на других молекулах РНК. Именно это свойство рибозимов данного класса позволило использовать их для создания высокоспецифичных эндорибонуклеаз на основе антисмысловых РНК, которые обладают способностью расщеплять in trans молекулы РНК, содержащие тринуклеотидные сайты расщепления. Для создания высокоспецифического рибозима, функционирующего по этому принципу, последовательность рибозима заключают в последовательность антисмысловой РНК таким образом, чтобы при образовании гибрида с РНК-мишенью в контакте с активным центром рибозима оказывалась тринуклеотидная последовательность сайта расщепления.