

Днк- и рнк-лигазы
Создание фосфодиэфирных связей в одноцепочечных разрывах двухцепочечной ДНК с помощью ДНК-лигаз является наряду с рестрикцией одним из важнейших этапов получения рекомбинантных ДНК in vitro. Наибольшее применение в генно-инженерных исследованиях находит ДНК-лигаза бактериофага Т4. Реакция лигирования протекает в два этапа (рис. II.3). Вначале образуется промежуточный комплекс фермент–АМР (этап 1), после чего остаток АМР переносится на 5’-фосфатную группу концевого нуклеотида в точке разрыва ДНК (этап 2). Образовавшаяся фосфодиэфирная связь гидролизуется во время нуклеофильной атаки 3’-ОН группы соседнего нуклеотида, что приводит к образованию новой фосфодиэфирной связи, восстанавливающей целостность сахаро-фосфатного остова ДНК. Т4-ДНК-лигаза осуществляет соединение фрагментов двухцепочечной ДНК, обладающих комплементарными "липкими" или "тупыми" концами. Как следует из механизма реакции, необходимым условием протекания лигирования является наличие 5’-концевого фосфата и 3'-концевого гидроксила в точках разрыва цепей ДНК. При этом эффективность соединения фрагментов ДНК по "тупым" концам Т4-ДНК-лигазой возрастает в присутствии Т4-РНК-лигазы, которая осуществляет ковалентное соединение 5’-фосфорилированных концов одноцепочечных ДНК или РНК с 3’-ОН группами одноцепочечных нуклеиновых кислот.
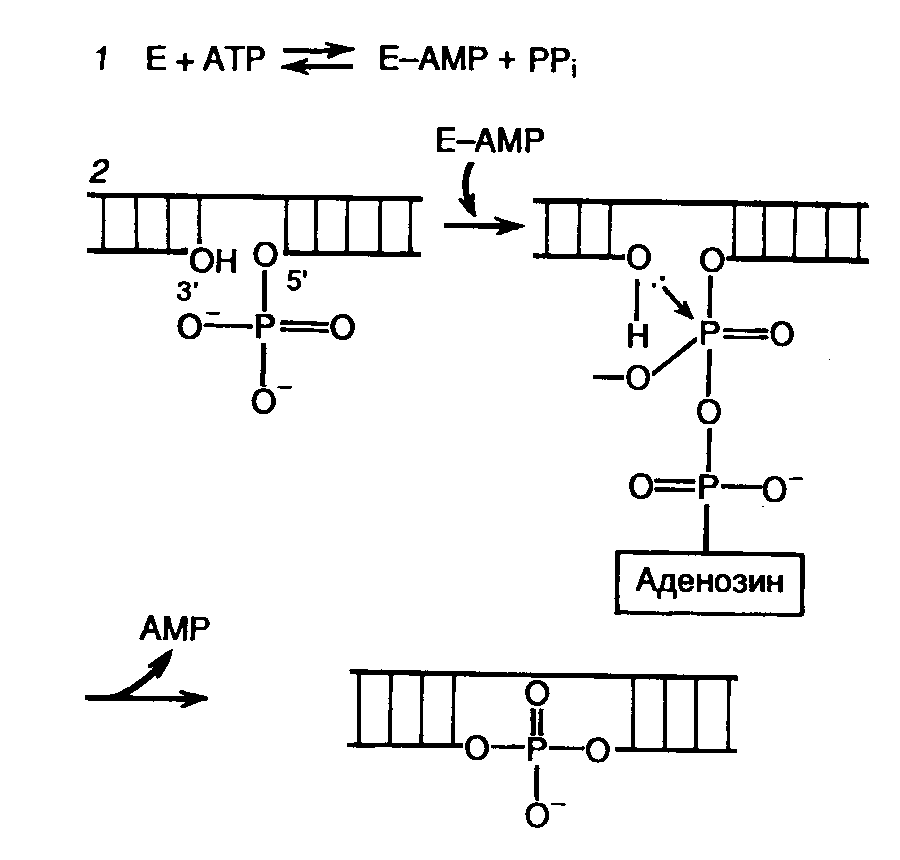
Рис. II.3. Механизм лигирования ДНК Т4-ДНК-лигазой
1 – образование промежуточного комплекса фермент Е–AMP; 2 – образование фосфодиэфирной связи
Ферменты матричного синтеза днк и рнк
К ферментам матричного синтеза нуклеиновых кислот относятся многочисленные ДНК- и РНК-зависимые ДНК- и РНК-полимеразы, осуществляющие зависимый от матричных ДНК или РНК синтез нуклеиновых кислот. Эти ферменты обычно используются в генной инженерии для получения двухцепочечных молекул ДНК из одноцепочечных, а также для обратной транскрипции, т.е. синтеза двухцепочечных ДНК, комплементарных мРНК, которые называют комплементарными ДНК (кДНК).
ДНК-зависимые ДНК-полимеразы. Среди ДНК-зависимых ДНК-полимераз наибольшее применение в генной инженерии находят ДНК-полимераза I E. coli и ее большой фрагмент (фрагмент Кленова), Т4-ДНК-полимераза и в последнее время термостабильные ДНК-полимеразы, особенно ДНК-полимераза Thermus aquaticus (Taq-полимераза). Все эти ферменты в присутствии ионов Mg2+ из четырех дезоксирибонуклеозидтрифосфатов (dATP, dCTP, dGTP и TTP) осуществляют синтез ДНК, комплементарной матричной ДНК, и для функционирования требуют наличия затравки на одноцепочечной матричной ДНК, т.е. олиго- или полидезоксирибонуклеотида со свободным 3’-ОН-концом, комплементарного матричной ДНК. ДНК-полимераза I E.coli состоит из одной полипептидной цепи с молекулярной массой около 109 кДа и обладает тремя активностями: полимеризующей в направлении 5’3’, 5’3’-экзонуклеазной и 3’5’-экзонуклеазной. Большой фрагмент ДНК-полимеразы I E. coli (фрагмент Кленова) является частью полипептидной цепи ДНК-полимеразы I с молекулярной массой около 76 кДа, у которой отсутствует домен, соответствующий 5’3’-экзонуклеазе. Как ДНК-полимераза I, так и ее фрагмент используются для введения радиоактивно меченных дезоксирибонуклеотидов в синтезируемые цепи ДНК путем ник-трансляции, т.е. перемещения одноцепочечного разрыва вдоль молекулы двухцепочечной ДНК, в котором 3’-ОН-конец используется в качестве затравки для ферментов. При этом ДНК-полимераза I прокладывает себе путь с помощью 5’3’-экзонуклеазы, а фрагмент Кленова вытесняет цепь ДНК с 5’-конца. Кроме того, фрагмент Кленова используют для синтеза второй цепи кДНК, секвенирования ДНК по методу Сенгера, заполнения 5’-выступающих "липких" концов ДНК с образованием "тупых" концов, введения концевой радиоактивной метки, а также для удаления 3’-выступающих концов рестрикционных фрагментов ДНК 3’5’-экзонуклеазой этого фермента. Как и ДНК-полимераза I E. coli, Т4-ДНК-полимераза обладает 3’5’- (но не 5’3’-) экзонуклеазной активностью, которая у последней, по крайней мере, в 200 раз выше. Это позволяет использовать Т4-ДНК-полимеразу, в частности, для введения радиоактивной метки путем реакции обмена немеченного 3’-концевого нуклеотида на меченный радиоактивным изотопом.
Полимеразная цепная реакция (ПЦР). Термостабильная Taq-полимераза в настоящее время широко используется для проведения полимеразной цепной реакции (ПЦР), а ее модифицированный аналог – и для секвенирования ДНК по методу Сенгера. Сущность ПЦР заключается в следующем (рис. II.4). Реакционная смесь обычно содержит исследуемую ДНК, dATP, dCTP, dGTP и TTP, Taq-полимеразу и два синтетических олигодезоксирибонуклеотидных праймера длиной 15–30 нуклеотидов, комплементарных участкам противоположных цепей ДНК, фланкирующих исследуемый участок ДНК-матрицы. ПЦР начинается с кратковременного (1–2 мин) прогревания реакционной смеси при 95o, в процессе которого происходит плавление цепей матричной ДНК и инактивация примесных белков, которые могут присутствовать в реакционной смеси, если используется недостаточно очищенная матричная ДНК, например из клинических образцов. Далее реакционную смесь охлаждают до температуры отжига праймеров с матричной ДНК (37–65o). В результате праймеры связываются с комплементарными им местами на матрице и с этого момента начинают служить затравками для синтеза Taq-полимеразой новых цепей ДНК, комплементарных каждой из цепей денатурированной матричной ДНК. Для проведения синтеза ДНК в оптимальных для Taq-полимеразы условиях температуру реакционной смеси поднимают до 72o и при такой температуре проводят элонгацию цепей вновь синтезирующейся ДНК в течение 0,5–2 мин. По окончании стадии синтеза ДНК заканчивается первый цикл ПЦР, после чего проводятся такие же второй, третий и т.д. в автоматическом режиме. По окончании третьего цикла уже образуется дискретный продукт ПЦР – фрагмент двухцепочечной ДНК, содержащий на своих 5’-концах последовательности нуклеотидов праймеров. Количество продукта ПЦР в реакционной смеси после завершения каждого цикла удваивается и, следовательно, по мере прохождения реакции экспоненциально возрастает, достигая нескольких микрограммов в реакционной смеси объемом 25–50 мкл. По окончании ПЦР содержимое реакционной смеси анализируют электрофорезом в агарозном геле в присутствии бромистого этидия, где продукт реакции выявляется в УФ-свете. Продукт ПЦР можно выделить из агарозного геля в чистом виде и работать с ним, как с обычным фрагментом двухцепочечной ДНК, т.е. гидролизовать рестриктазами, клонировать в плазмидных векторах по "липким" и "тупым" концам, секвенировать или использовать в качестве зондов при гибридизации.
С помощью ПЦР можно амплифицировать in vitro сегменты ДНК длиной от 0,1 до 5–7 т.п.о. и более, а для получения положительного результата достаточно присутствия в реакционной смеси одной–двух копий амплифицируемой последовательности нуклеотидов (например геномной ДНК, содержащейся в одной–двух соматических клетках). При этом не возникает необходимости в тщательной очистке матрицы, так как большинство попадающих в реакционную смесь белков и ферментов инактивируется в первых же циклах ПЦР и не оказывает влияния на протекание реакции при высоких температурах. Благодаря таким особенностям, как простота постановки, высокая чувствительность и воспроизводимость, ПЦР в последнее время получила широкое распространение в фундаментальных и прикладных исследованиях по молекулярной биологии и генетике. Возможность амплификации любого сегмента ДНК, последовательность нуклеотидов которого известна, и получение его по завершении ПЦР в гомогенном виде и препаративном количестве делают ПЦР альтернативным методом молекулярного клонирования коротких фрагментов ДНК. При этом не возникает необходимости в применении тех сложных методических приемов, которые используют в генной инженерии при обычном клонировании.
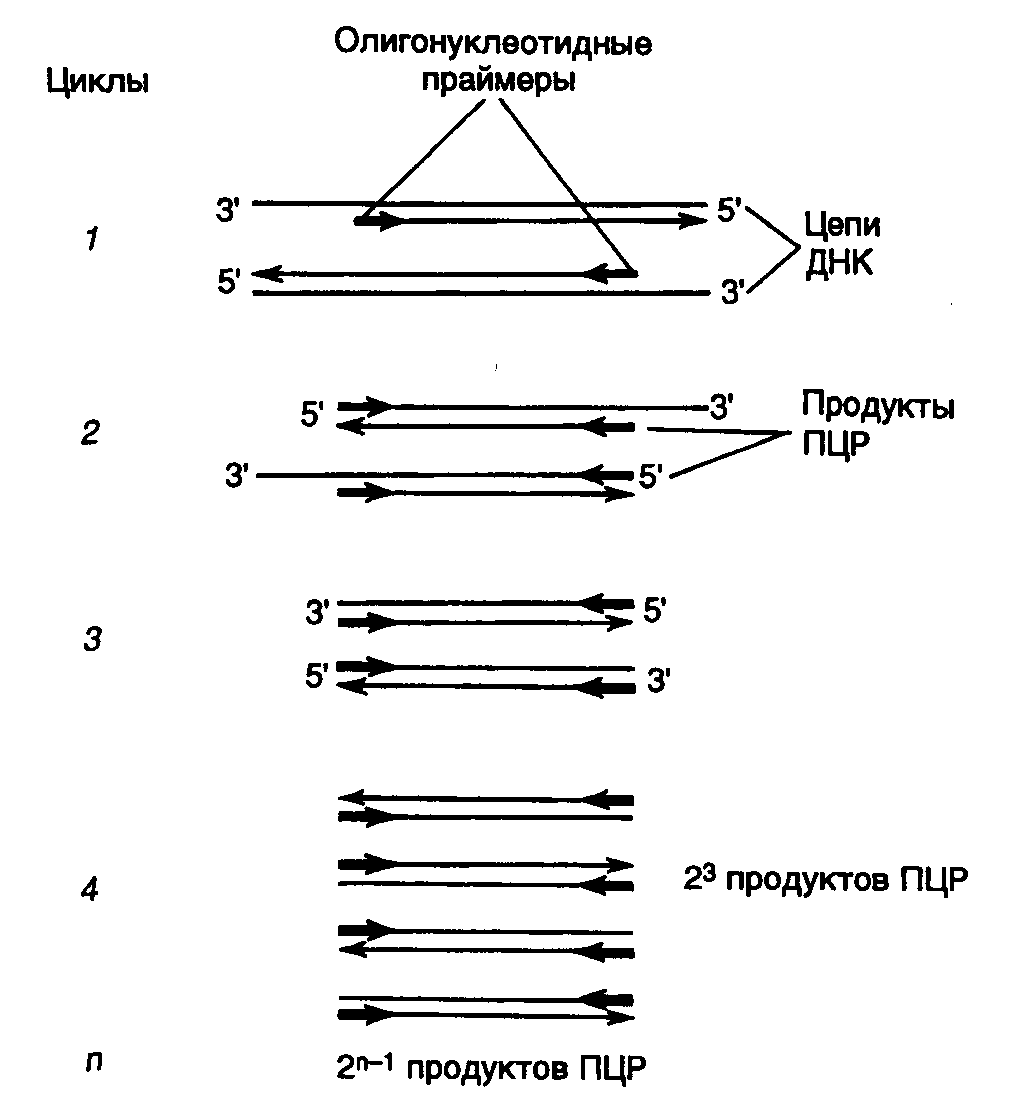
Рис. II.4. Полимеразная цепная реакция (ПЦР)
Поскольку специфичность ПЦР зависит главным образом от соответствия последовательности нуклеотидов праймеров первичной структуре амплифицируемых фрагментов ДНК, эта специфичность приближается к абсолютной при повышении длины праймеров до 17–20 нуклеотидов. Такая высокая специфичность и чувствительность позволяют использовать ПЦР для ДНК-диагностики инфекционных и наследственных заболеваний человека, а также вирусоносительства в тех случаях, когда вирусом в латентной форме заражены лишь отдельные соматические клетки, например в случае вируса иммунодефицита человека (ВИЧ) (см. главу 11). Разработка метода ПЦР во многом расширила методические возможности молекулярной генетики, и в частности генной инженерии, причем настолько, что это кардинально изменило и усилило исследовательский потенциал многих ее направлений.
Точность амплификации ДНК термостабильными ДНК-полимеразами является особенно критическим параметром в том случае, если продукты ПЦР используются далее для конструирования экспрессирующихся генов. Необходимость в уменьшении частоты мутаций стимулирует поиски новых термостабильных ДНК-полимераз, амплифицирующих ДНК с большей точностью. В табл. II.2 приведены результаты испытаний точности синтеза ДНК различными коммерческими ДНК-полимеразами.
Относительно высокая точность синтеза ДНК in vitro ДНК-полимеразами Pfu и Vent обусловлена наличием у них 3’-5’-экзонуклеазной корректирующей активности. Частота ошибочно включенных нуклеотидов в результате одновременного функционирования двух разных ДНК-полимераз в смесях, применяемых для амплификации длинных сегментов ДНК (long PCR), оказывается ниже, чем при использовании Taq-полимеразы, но выше, чем для очищенной ДНК-полимеразы Pfu.
При проведении аллель-специфической ПЦР (см. раздел 11.1) необходимо учитывать способность Taq-полимеразы инициировать синтез ДНК на праймерах, 3’-концевой нуклеотид которых некомплементарен матричной ДНК. При исследовании этого вопроса оказалось, что не все пары
Таблица II.2