

Идентификация личности на основе минисателлитной днк: определение отцовства
Определение отцовства представляет собой серьезную социальную, юридическую и медицинскую проблему. Решение этой задачи часто требуется в судах, при разрешении частных споров, для пренатальной (внутриутробной) диагностики, генетических консультаций и при пересадках органов. Например, только в США в 1990 г. было проведено более 120000 тестов на определение отцовства, и это число быстро возрастает. Мировой рынок таких диагностических тест-систем оценивался в 1994 г. более чем в 1 млрд долларов и сейчас является крупнейшим рынком среди молекулярно-генетических диагностикумов.
С использованием диагностических тест-систем на основе минисателлитных ДНК определение отцовства получило прочную научную основу. С этой целью в настоящее время используют два подхода. При одном из них применяют олигонуклеотидные зонды, специфичные в отношении многих минисателлитных локусов, при другом – наборы зондов (или праймеров), специфичных в отношении отдельных полиморфных локусов VNTR (подробнее о VNTR см. раздел 1.3.1).
Теоретические аспекты возможности определения отцовства. Возможность определения отцовства, так же как и отнесение биологических образцов, содержащих ДНК, к тому или иному человеку, основано на наличии в геноме человека четких генетических маркеров в виде определенных последовательностей ДНК, набор которых уникален для конкретного индивидуума. Теоретически такими маркерами могли бы быть любые последовательности нуклеотидов ДНК, для которых характерна большая изменчивость в популяциях человека. Как было уже отмечено выше, тандемно повторяющиеся последовательности минисателлитов (VNTR) являются одними из наиболее полиморфных последовательностей нуклеотидов в геноме человека. Поэтому неудивительно, что именно они были использованы для идентификации личности. Наличие сочетания генетических маркеров среди VNTR, общих для мужчины, женщины и спорного ребенка, в ряде случаев может однозначно указывать на родственные связи между ними.
Отсутствие общих генетических маркеров у обследуемых ребенка и мужчины однозначно исключает последнего как отца ребенка. Однако обнаружение у них общих маркеров еще не может быть доказательством того, что подозреваемый мужчина является отцом. Доказательства отцовства основываются на простых статистических расчетах, в которых учитываются частоты встречаемости в популяции общих аллелей исследуемых VNTR-локусов. Рассчитывается отношение (X/Y) вероятности (X) получения наблюдаемого набора маркеров возможного настоящего отца к вероятности (Y) обнаружения этого набора маркеров у любого, выбранного наугад человека, принадлежащего этой популяции. Такое отношение вероятностей получило название индекса отцовства (paternity index – PI).
При расчетах PI возникает еще одна проблема, связанная с определением вероятности (X) того, что обследуемый мужчина является отцом. Значение такой вероятности необходимо иметь до проведения каких-либо диагностических тестов. Обычно используемое значение 0,5 нельзя считать достаточно обоснованным во всех конкретных случаях. К счастью, при очень больших значениях PI (>104), которые обычно получаются при таких исследованиях ДНК, выбор этой исходной вероятности оказывается практически несущественным. Как правило, это получается вследствие низких значений вероятности Y.
Определение отцовства с использованием олигонуклеотидных зондов, специфичных в отношении нескольких VNTR-локусов. В 1985 г. И.О. Джеффрисом и соавторами впервые было показано, что олигонуклеотидные зонды, комплементарные последовательностям миоглобинового гена человека, одновременно обладают способностью гибридизоваться по Саузерну с множественными локусами минисателлитной ДНК. Профили гибридизации оказались специфичными для отдельных индивидуумов. Совокупность электрофоретически разделяющихся рестрикционных фрагментов анализируемой ДНК, выявляемых после проведения гибридизации с мечеными зондами, которые специфичны в отношении полиморфных минисателлитных локусов, получила название ДНК-фингерпринтов, или генетических отпечатков пальцев. С помощью таких и других аналогичных олигонуклеотидных зондов удается выявлять на одной электрофореграмме до 15–20 различных фрагментов ДНК одного индивидуума, молекулярная масса которых превышает 3,5 т.п.о., а также много более мелких фрагментов, которые не учитываются при определении отцовства этим методом.

Рис. II.36. Примеры исключения и доказательства отцовства с помощью ДНК-типирования
Результаты гибридизации по Саузерну с зондом F10 показывают полную идентичность фрагментов ДНК у ребенка и отца (дорожки 2 и 3), что рассматривается как доказательство отцовства, либо выявляют, по крайней мере, 6 дополнительных фрагментов ДНК у ребенка, обозначенных стрелками, которые отсутствуют у отца (дорожки 5 и 6 – исключение отцовства)
На рис. II.36 показаны результаты одного из таких опытов. Интерпретация ДНК-фингерпринтов, полученных при анализе множественных локусов, основана на трех постулатах. Прежде всего, предполагается, что фрагменты ДНК, видимые на фингерпринтах, являются аллельными продуктами отдельных генетических локусов человека и передаются потомству независимо друг от друга. Во-вторых, считается, что для каждого генетического локуса частоты встречаемости в популяции отдельных аллелей следуют нормальному распределению Пуассона. И, наконец, принимается, что фрагменты ДНК, электрофоретическая подвижность которых совпадает, представляют один и тот же аллель конкретного локуса. Накопленный опыт работы с ДНК-фингерпринтами показывает, что первое допущение соблюдается достаточно хорошо: аллелизм (парность гомологичных генов, определяющих разные фенотипические признаки у диплоидных организмов) и генетическое сцепление между исследуемыми локусами наблюдаются редко. Невыполнение второго предположения не сказывается серьезно на результатах тестирования, поскольку выводы делаются без учета частоты встречаемости отдельного аллеля на основе совпадения структуры (фрагментов ДНК) многих локусов. Третий постулат является более спорным, однако его применение придает значениям индекса отцовства стабильность.
С этими исходными условиями статистическая оценка ДНК-фингерпринтов множественных локусов основывается только на одном параметре: средней доле фрагментов ДНК (x), которые совпадают у людей без родственных связей. Такой параметр в большей степени зависит от техники лабораторных исследований, чем от свойств обследуемой популяции. Это прежде всего способность используемой системы к электрофоретическому разделению индивидуальных фрагментов ДНК (т.е. разрешающая способность используемого метода), принципы выбора конкретных фрагментов ДНК для анализа, а также критерии принятия решения об идентичности сравниваемых фрагментов ДНК. Следовательно, параметр x может варьировать при сравнении результатов, получаемых в разных лабораториях, но эти различия будут постоянно сохраняться для различных популяций и субпопуляций. Действительно, при использовании, например зондов 33.6 и 33.15, оказалось, что x один и тот же у неродственных индивидуумов, в парах муж–жена и в различных этнических группах.
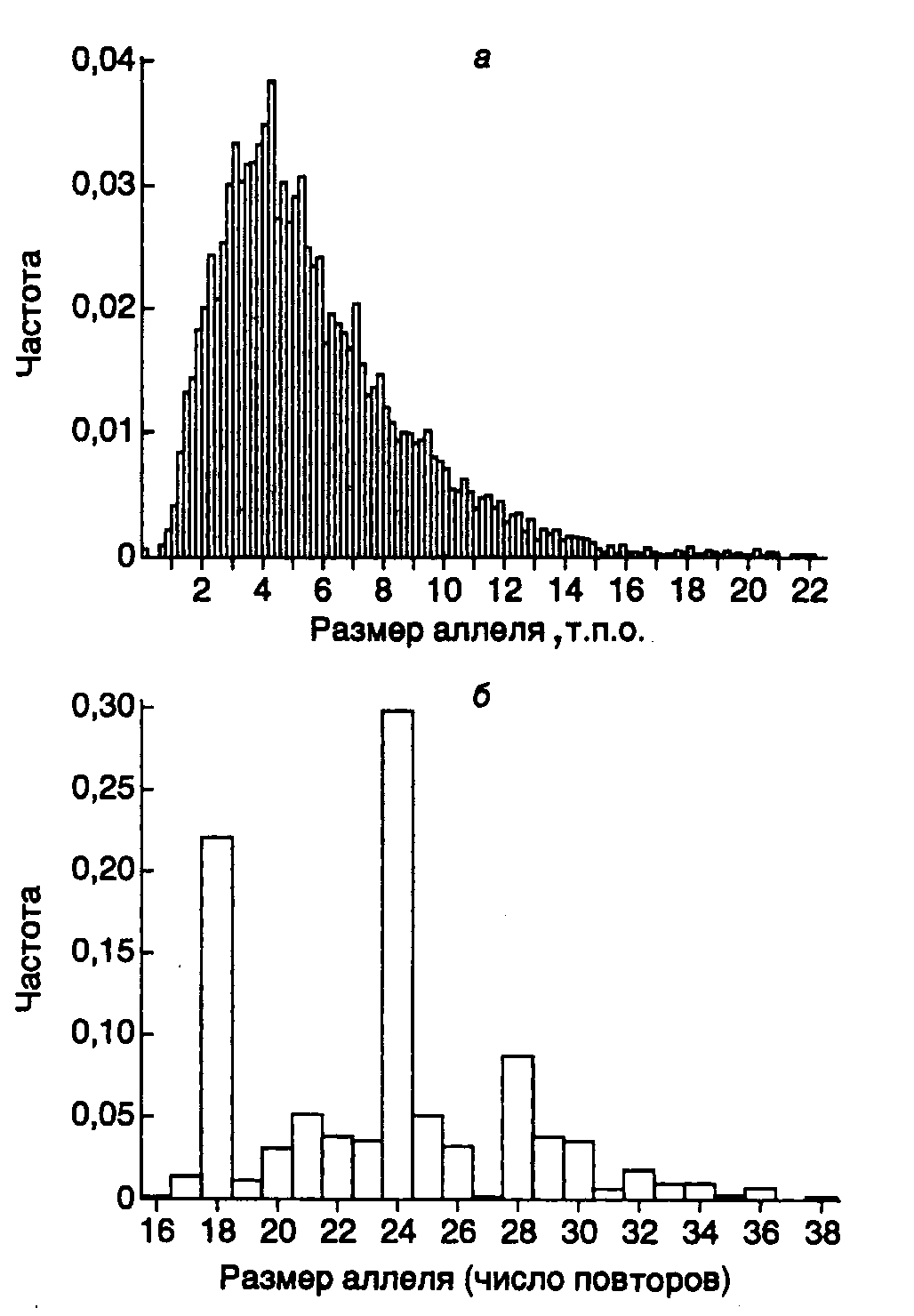
Рис. II.37. Сравнение информативности двух минисателлитных зондов при идентификации личности
Частоты встречаемости минисателлитных локусов D1S7 (а) и D1S80 (б) определенного размера в популяции оценивали гибридизацией по Саузерну после расщепления ДНК рестриктазойHaeIII (а) или ПЦР (б). Для локуса D1S7 характерно квази-непрерывное унимодальное распределение, тогда как локус D1S80 характеризуется меньшей гетерозиготностью (84%) с небольшим числом дискретных аллелей, для которых характерно мультимодальное распределение
Определение отцовства с использованием ДНК-зондов, специфичных в отношении только одного локуса. ДНК-фингерпринты, получаемые при одновременном исследовании многих локусов, отражают скорее фенотип индивидуума, чем его генотип. Действительно, получаемая в итоге картина заключает в себе множество полос ДНК, в том числе и неразделившиеся, а также слабо разделившиеся фракции. Такая электрофореграмма напоминает сложный фенотипический признак, например форму лица человека, которая является результатом экспрессии громадного числа генов. В отличие от этого с помощью ДНК-зондов, представляющих собой клонированные последовательности минисателлитов и взаимодействующих только с одним локусом, можно получать истинную информацию о генотипе изучаемого организма. Таким образом, имея дело с отдельными полиморфными локусами человека, исследователи получают в свои руки систему кодоминантных аллелей (т.е. аллелей, совместно участвующих в формировании фенотипических признаков), наследуемых по законам Менделя. Именно понимание наследования таких минисателлитных локусов и привело к широкому распространению однолокусного метода определения отцовства.
В настоящее время получены сотни клонов минисателлитной ДНК и на их основе разработаны комбинации зондов, пригодные для определения отцовства. При выборе зондов для таких минисателлитных локусов обычно руководствуются следующими критериями: зонды должны обладать строгой локус-специфичностью, а тестируемые локусы быть несцепленными (передаваться потомству независимо друг от друга) и обладать достаточной, но не чрезмерной генетической стабильностью. В частности, среди наиболее широко используемых зондов MS1 (D1S7) соответствует генетическому локусу, гетерозиготному в 99% случаев, однако он мутирует с очень высокой частотой (0,05 на гамету) и поэтому, несмотря на высокую информативность, не используется при определении отцовства (рис. II.37,а). В то же время для локуса D1S80 со значительно меньшей вариабельностью (84% гетерозигот) характерно образование кластеров в частотах распределения фрагмента ДНК по длине (см. рис. II.37,б). Поэтому небольшие ошибки в определении длины аллелей могут привести к значительным ошибкам в оценке частоты их встречаемости. Такие локусы достаточно легко изменяются в результате генетического дрейфа и инбридинга.
В настоящее время разработана теория, указывающая на то, сколько локусов должно быть типировано для получения правильного ответа об отцовстве при известных значениях гетерозиготности этих локусов в популяции. Например, если используются локусы, гетерозиготность которых составляет 90%, для установления отцовства необходимо проанализировать шесть таких локусов. В США в настоящее время для этих целей обычно используется набор из трех–пяти однолокусных зондов. Однолокусные зонды обладают низкой разрешающей способностью относительно братьев и сестер (сибсов). В частности, с помощью одного такого зонда можно лишь с вероятностью 75% обнаружить генетические различия между сибсами, и эта вероятность увеличивается до 99,6% при использовании четырех зондов. Данный факт приобретает особую важность, когда при определении отцовства необходимо сделать выбор между братьями.
Особенности определения отцовства по отдельным локусам с использованием ПЦР. В качестве альтернативы однолокусным зондам в последнее время для определения отцовства часто используют ПЦР. Отдельные мини- и микросателлитные локусы могут быть амплифицированы с помощью праймеров, комплементарных уникальным последовательностям ДНК, фланкирующим эти повторяющиеся последовательности. При идентификации личности метод ПЦР, который по своей сути является одной из разновидностей однолокусной методики, поскольку имеет дело с отдельными локусами, обладает, по крайней мере, двумя преимуществами перед однолокусными зондами. Во-первых, популяционный полиморфизм длин ДНК аллелей, исследуемых с использованием этого метода, носит более дискретный характер, чем у аллелей, изучаемых с помощью однолокусных зондов (см. рис. II.37,а,б). Это обстоятельство облегчает последующее вычисление индекса отцовства. Во-вторых, метод ПЦР обладает гораздо большей чувствительностью и может быть использован при анализе образцов, содержащих <1 нг геномной ДНК и полученных из разных источников (см. раздел 11.1.1).
К недостаткам ПЦР в применении к определению отцовства следует отнести низкую информативность полиморфных микросателлитов и коротких минисателлитов. Это связано с тем, что они обладают <90% гетерозиготности, небольшим числом аллелей и имеют тенденцию к образованию кластеров по размерам (см. рис. II.37,б). Кроме того, на распределение таких последовательностей в геноме оказывают влияние инбридинг и принадлежность индивидуумов к определенным этническим группам. Количество локусов минисателлитов, которое необходимо исследовать методом ПЦР для определения отцовства, приближается к 11, а микросателлитов – к 18.
Для типирования с помощью ПЦР наиболее часто используются три минисателлитных локуса человека: в гене аполипопротеина B (APOB), D17S5 (известный также как локус D17S30) и D1S80. Все три локуса легко амплифицируются (максимальный размер их аллельных вариантов не превышает 1 т.п.о.) и легко обнаруживаются с помощью электрофореза. Однако для них характерны низкий уровень гетерозиготности и малая изменчивость (что выражается в небольшом числе известных аллелей). Мутации в этих микросателлитах возникают очень редко.
Одним из путей повышения информативности полиморфизмов микросателлитов при ДНК-типировании является одновременная амплификация двух тесно сцепленных микросателлитных локусов, сочетания которых формируют множество гаплотипов. Например, одновременная амплификация двух GATA-повторов, локализованных в интроне 40 гена фактора фон Виллебранда, которые разделены последовательностью длиной в 212 п.о., обнаружила суммарный уровень гетерозиготности объединенного локуса 93%. При этом уровни гетерозиготности индивидуальных локусов составляли лишь 72 и 78% соответственно.
В заключение необходимо еще раз отметить, что по своей логике современные методы определения отцовства, основанные на ДНК-типировании, несколько противоречивы. Если отрицательное заключение об отцовстве, основанное на несовпадении аллелей анализируемых мини- или микросателлитных локусов, абсолютно и не подлежит сомнению, то положительный вывод может быть сделан лишь с некоторой долей вероятности, которая основана на частоте встречаемости конкретных аллелей анализируемых локусов в популяции. С другой стороны, в результате методических ошибок легко могут быть сделаны ложноотрицательные выводы, однако положительное заключение об отцовстве в результате лабораторной методической ошибки практически исключено.
Высокая информативность многолокусных ДНК-фингерпринтов подтверждена большим числом генетических и популяционных исследований. Эмпирические данные, полученные при обследовании тысяч семей, показали, что с помощью многолокусных зондов можно разрешать все спорные случаи отцовства. Использование однолокусных зондов затруднено невозможностью создания полной классификации соответствующих аллелей в популяции из-за кажущегося непрерывного распределения их по размерам. Однако и в этом случае на практике проблема полностью решается с помощью набора из пяти–шести однолокусных зондов. Применение ПЦР ограничивается большой эволюционной консервативностью амплифицируемых мини- и микросателлитных локусов и, как следствие, малым суммарным числом аллелей. Однако ПЦР бывает очень полезна на первых этапах исследования из-за методической простоты постановки опытов, особенно в условиях малой доступности исходного биологического материала. Все три группы методов хорошо дополняют друг друга и в разных сочетаниях в спорных случаях позволяют однозначно идентифицировать личность человека.