

Передача сигнала и вторичные мессенджеры
Жизнь любой клетки, включая глобальные процессы ее роста, деления и даже гибели, зависит от внешних регуляторных сигналов, которые она воспринимает. Такими сигналами могут быть физические воздействия (температура, ионизирующее и другое электромагнитное излучение) или многочисленные химические соединения. Хорошо изученными веществами, которые организм использует для регуляции жизнедеятельности клеток, являются, например стероидные гормоны, цитокины или факторы роста, которые, достигая клеток-мишеней, вызывают в них специфические метаболические изменения, связанные в том числе и с изменением экспрессии больших групп генов. Не менее сильный и часто также специфический ответ вызывают различные физиологически активные вещества экзогенного происхождения, например феромоны или токсины.
Все эти сигналы, передающиеся через соответствующие сигнальные молекулы, являются первичными по отношению к тем каскадам биохимических реакций, которые запускаются в клетках в ответ на их воздействие. Первичные сигналы распознаются клетками благодаря наличию у них специальных молекул-рецепторов белковой природы, взаимодействующих с первичными сигнальными молекулами или воздействиями физической природы. Первичный сигнал, как правило, не действует прямо на те метаболические процессы в клетке, для регуляции которых он предназначен. Вместо этого воспринимающий его рецептор инициирует образование в клетке промежуточных химических соединений, запускающих внутриклеточные процессы, воздействие на которые было целью первичного внеклеточного сигнала. Поскольку такие промежуточные соединения несут в себе информацию о первичном регуляторном сигнале и являются вторичными его переносчиками, они получили название вторичных мессенджеров. Ими могут быть различные ионы, циклические нуклеотиды, продукты деградации липидов и целый ряд других химических соединений биогенного происхождения.
Использование эукариотами системы вторичных мессенджеров переводит их на новый уровень интеграции всех метаболических и катаболических процессов, что необходимо для существования многоклеточных организмов. В частности, вторичные мессенджеры позволяют многократно усиливать первичный регуляторный сигнал от внеклеточных регуляторных молекул, которые благодаря этому осуществляют свое действие, находясь в небольших концентрациях во внеклеточном пространстве. Кроме того, многие группы клеток и тканей приобретают способность к однотипной и одновременной реакции на первичный регуляторный сигнал, например на действие гормона какого-либо органа эндокринной системы. Это обеспечивает возможность быстрой адаптации многоклеточного организма к изменяющимся условиям внутренней и окружающей среды.
Трансмембранный перенос первичных сигналов. Для того чтобы первичный регуляторный сигнал достиг ядра и оказал свое воздействие на экспрессию генов-мишеней, он должен пройти через двухслойную мембрану именно тех клеток, которым он предназначен. Как правило, это достигается благодаря наличию на поверхности клеток рецепторов белковой природы, специфически выбирающих из окружающей среды сигналы, распознать которые они в состоянии (рис. I.22). В простейшем случае, когда в качестве низкомолекулярных регуляторов выступают гидрофобные химические соединения, растворимые в липидах мембран (например стероидные гормоны), для их переноса не используются рецепторы, и они проникают в клетку путем радиальной диффузии. Внутри клеток такие соединения специфически взаимодействуют с белковыми рецепторами, а образующийся комплекс переносится в ядро, где оказывает свое регуляторное воздействие на транскрипцию соответствующих генов (см. рис. I.22,а).
В отличие от этого рецепторы мембран, ориентированные во внеклеточное пространство, обладают способностью осуществлять транспорт лиганда-регулятора внутрь клеток посредством эндоцитоза (поглощения путем втягивания мембраны) комплекса лиганд-рецептор в составе мембранных везикул. Такой механизм используется, в частности, для переноса внутрь клеток молекул холестерина, ассоциированных с рецепторами липопротеинов низкой плотности (см. рис. I.22,б). Другой тип рецепторов, ориентированных на внеклеточные лиганды, – это трансмембранные молекулы или группа молекул. Взаимодействие с лигандом внешней части таких молекул сопровождается индукцией ферментативной активности, ассоциированной с внутриклеточной частью того же самого полипептида (см. рис. I.22,в). Примерами подобных рецепторов, обладающих активностью тирозиновых протеинкиназ, являются рецепторы инсулина, эпидермального фактора роста или фактора роста тромбоцитов. В синапсах нейронов и местах контакта нейромышечных тканей лиганды-нейромедиаторы (например ацетилхолин или -аминомасляная кислота) взаимодействуют с трансмембранными ионными каналами (см. рис. I.22,г). В ответ на это происходит открытие ионных каналов, сопровождаемое перемещением ионов через мембрану и быстрым изменением трансмембранного электрического потенциала. Другие трансмембранные рецепторы осуществляют связь белков внеклеточного матрикса с микрофиламентами цитоскелета клеток и регуляцию формы клеток, зависящую от внеклеточного матрикса, их подвижности и роста (см. рис. I.22,д). Наконец, большая группа внеклеточных сигналов распознается рецепторами, ассоциированными на внутренней поверхности мембраны с GTP-связывающими белками, которые, в свою очередь, в ответ на первичный сигнал начинают синтез вторичных мессенджеров, регулирующих активность внутриклеточных белков (см. рис. I.22,е). Классификация по структурному признаку рецепторов, осуществляющих перенос сигнала в клетки через мембраны, приведена в табл. I.12.
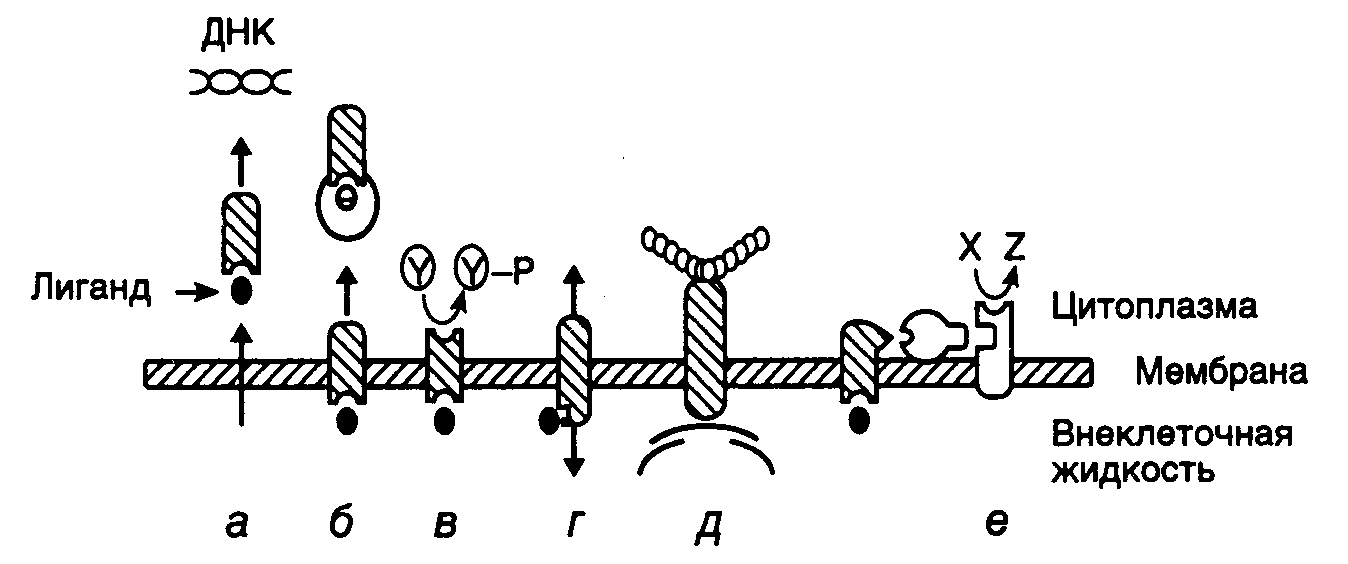
Рис. I.22. Способы передачи внеклеточных регуляторных сигналов через мембраны эукариотических клеток (а–е)
Y и Y–P – нефосфорилированные и фосфорилированные остатки Tyr в белках соответственно. Показано также превращение предшественника X во вторичный мессенджер Z
Все рецепторы, участвующие в трансмембранной передаче сигнала, подразделяют на три класса. При этом, как правило, учитывается сходство или различие вторичных структур субъединиц, а не особенности их аминокислотных последовательностей. Рецепторы 1-го класса образуют олигомерные структуры вокруг пор в мембранах. Перенос сигнала в этом случае происходит в результате открытия или (в одном случае) закрытия ионных каналов. Основная часть рецепторов 2-го класса погружена в мембраны, и каждая из субъединиц содержит последовательности, распознаваемые G-белками (см. ниже). Для всех субъединиц этого класса характерно наличие трансмембранной (ТМ) последовательности, которая 7 раз пересекает мембрану. Субъединицы рецепторов 3-го класса минимально погружены в мембраны, что обеспечивает подвижность рецепторов и возможность их интернализации (перехода в цитоплазму клеток в составе мембранной везикулы). Большая часть полипептидных цепей этих субъединиц экспонирована наружу клеток.
Вторичные мессенджеры. Гипотеза о том, что действие гормонов на метаболизм клеток и экспрессию генов опосредуется внутриклеточными вторичными мессенджерами, впервые появилась после открытия в конце 1950-х годов Е. Сазерлендом циклического аденозин-3’,5’-монофосфата (cAMP). К настоящему времени список вторичных мессенджеров расширился и включает циклический гуанозин-3’,5’-монофосфат, фосфоинозитиды, ионы Ca2+ и H+, метаболиты ретиноевой и арахидоновой кислот, закись азота (NO), а также некоторые другие химические соединения биогенного происхождения. Подробное рассмотрение особенностей механизма действия каждого из них выходит за рамки данной монографии, хотя все они могут оказывать специфическое влияние на экспрессию генов.
Таблица I.12