

Этапы транскрипции
Процесс транскрипции в настоящее время принято подразделять на 4 основные стадии: 1) связывание молекул РНК-полимеразы с ДНК и распознавание промотора; 2) инициация; 3) элонгация; 4) терминация. Как будет видно из дальнейшего изложения, три последних этапа характерны для биосинтеза большинства других макромолекул клетки, особенно для тех из них, синтез которых является матричным, в частности белков. После связывания с ДНК молекулы РНК-полимеразы осуществляют поиск промоторов, на которых происходит формирование инициационных комплексов. Начальная стадия инициации транскрипции завершается образованием нескольких первых фосфодиэфирных связей в молекуле вновь синтезируемой РНК, после чего транскрипция переходит в стадию элонгации – последовательного удлинения синтезируемых молекул РНК. Стадия элонгации заканчивается по достижении молекулами РНК-полимераз специальных регуляторных последовательностей ДНК, называемых терминаторами транскрипции, после чего происходит освобождение синтезированных молекул РНК и РНК-полимераз из транскрипционных комплексов. Освободившиеся молекулы РНК-полимераз приобретают способность вступать в новый цикл транскрипции. Следует помнить, что четкое разделение единого процесса транскрипции на отдельные стадии условно; оно используется главным образом для удобства описания механизмов биосинтеза РНК и является упрощенной моделью. Основные этапы транскрипции, а также дальнейшие пути реализации генетической информации схематически представлены на рис. I.6.
В обычных условиях холофермент РНК-полимераз эубактерий для инициации транскрипции не требует дополнительных факторов. В отличие от этого для точной инициации транскрипции РНК-полимеразой II требуется наличие, кроме ее субъединиц, еще и основных факторов транскрипции. Синтез РНК, который не зависит от присутствия регуляторных молекул, получил название базальной транскрипции. Транскрипция в клетках является регулируемым процессом, который, как уже упоминалось выше, требует участия белков-активаторов или репрессоров. Белок-активатор (тканеспецифический фактор транскрипции) взаимодействует с регуляторными последовательностями ДНК и активирует синтез РНК. Такая транскрипция получила название индуцированной, или активированной. Следовательно, базальная транскрипция не может происходить in vivo, и этот термин используется только при описании результатов исследований синтеза РНК in vitro, в бесклеточных системах транскрипции.
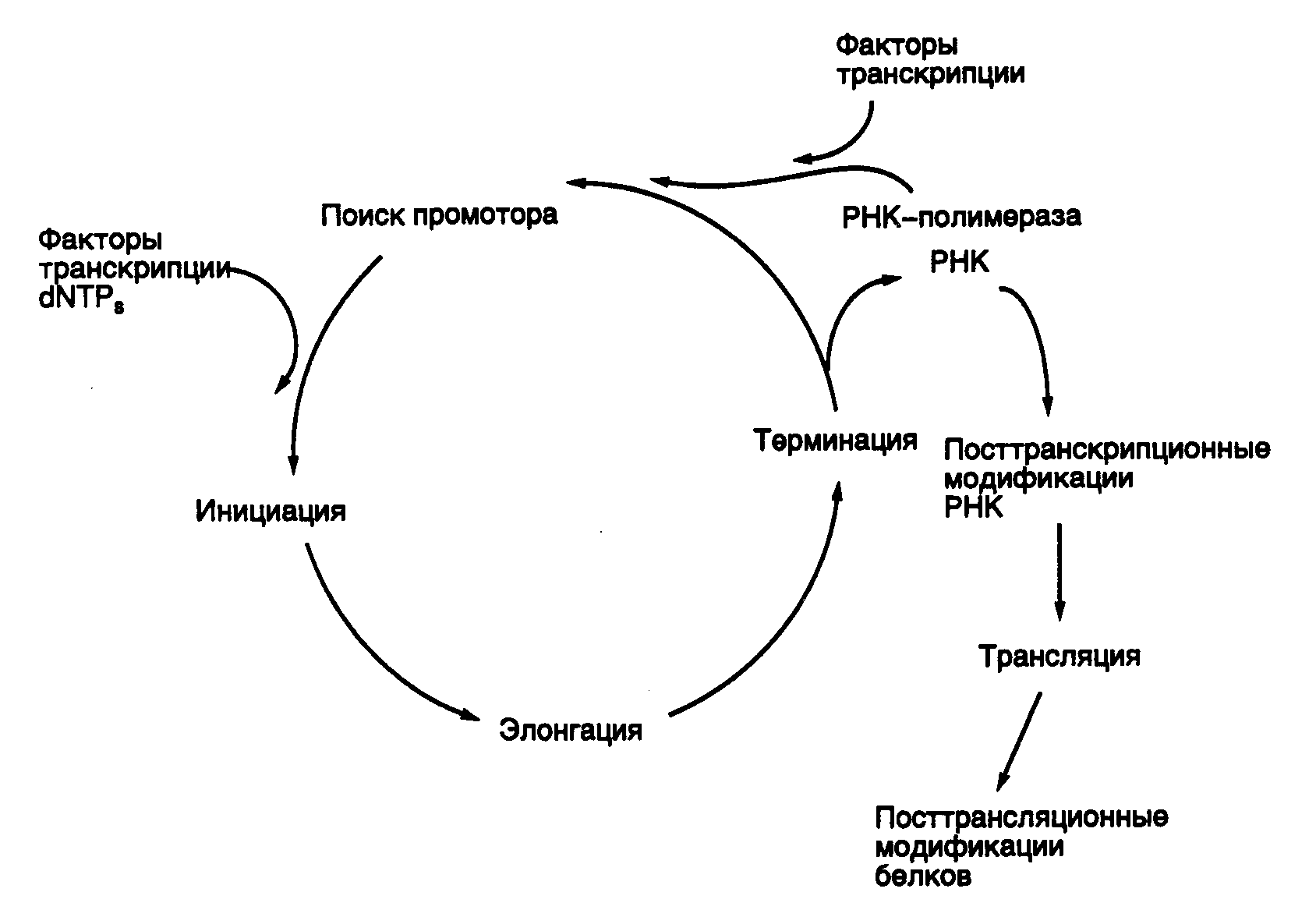
Рис. I.6. Цикл транскрипции и последующие пути реализации генетической информации
Связывание молекул РНК-полимеразы с ДНК и поиск промоторов. Механизм поиска промоторов на ДНК молекулами РНК-полимеразы в настоящее время до конца не выяснен. Принято считать, что после первоначального непрочного связывания с ДНК в случайных местах молекулы РНК-полимеразы перемещаются вдоль двойной спирали ДНК до тех пор, пока не обнаруживают последовательности нуклеотидов промоторов, на которых взаимодействие фермента с ДНК становится более прочным. Во время движения молекулы РНК-полимеразы могут периодически отделяться от ДНК и связываться с ней на новом месте, что ускоряет процесс поиска промоторов. Как уже упоминалось выше, в связывании с ДНК участвует -субъединица РНК-полимеразы E. coli, а - и особенно -субъединицы необходимы для специфического распознавания промоторов. Установлено, что холофермент РНК-полимеразы E. coli (минимальный фермент, содержащий -субъединицу) закрывает в области промотора участок ДНК длиной ~50 п.о. При этом -субъединицы контактируют с ДНК в области 35-го нуклеотида промотора.
Инициация транскрипции. Инициация транскрипции начинается со сборки на промоторе прединициационного комплекса, в состав которого входят молекулы РНК-полимеразы и матричной ДНК. Если в случае РНК-полимеразы E. coli и других прокариот для осуществления этого процесса нет необходимости в присутствии других белковых факторов, то механизм сборки инициационного комплекса с участием РНК-полимеразы II носит более сложный характер. В настоящее время существуют две модели инициации транскрипции РНК-полимеразой II. В соответствии с одной из них на промоторе происходит постепенная (ступенчатая) сборка инициационного комплекса из отдельных компонентов. Другая модель акцентирует внимание на то, что Pol II может входить в состав инициационного комплекса в виде холофермента, состоящего из многих субъединиц. Хотя вторая модель становится доминирующей, ниже будет подробнее рассмотрена первая модель, более наглядно описывающая процесс инициации транскрипции у эукариот.
Сборка такого комплекса начинается с последовательного связывания с промотором основных факторов транскрипции (табл. I.4). Обычно факторами транскрипции называют белки или белковые комплексы, непосредственно не участвующие в каталитическом акте образования РНК, но необходимые для прохождения основных этапов транскрипции и ее регуляции. По функциональному признаку принято различать три класса факторов транскрипции. К первому классу относятся основные факторы транскрипции, обеспечивающие нерегулируемый базальный уровень транскрипции и функционирующие в клетках всех типов.
Таблица I.4