

Физические карты низкого разрешения
В отличие от рассмотренных выше генетических карт сцепления физические карты генома отражают реальное расстояние между маркерами, выражаемое в парах оснований. Физические карты различаются по степени их разрешения, т.е. по тем деталям структуры генома, которые на них представлены (рис. II.39). Исчерпывающая физическая карта генома человека максимального разрешения будет содержать полную нуклеотидную последовательность всех его хромосом. На другом полюсе физических карт с минимальным разрешением находятся хромосомные (цитогенетические) карты генома.

Рис. II.39. Четыре типа генетических карт геномной ДНК и их взаимоотношения
1 – генетическая карта сцепления, 2 – физическая рестрикционная карта, пробелы обозначают места расщепления ДНК рестриктазами, 3 – физическая карта контигов, показаны перекрывающиеся клоны ДНК, полученные с помощью YAC-векторов, 4 – исчерпывающая физическая карта в виде последовательности нуклеотидов ДНК. На всех картах представлен один и тот же участок хромосомы
Хромосомные карты. Хромосомные карты генома человека получают локализацией генетических маркеров на индивидуальных хромосомах с использованием цитогенетических методов, включая авторадиографию и FISH. В последних двух случаях радиоактивная или флуоресцентная метки, ассоциированные с исследуемыми генетическими локусами интактных хромосом, выявляются с помощью световой микроскопии. Еще совсем недавно хромосомные карты позволяли локализовать исследуемый фрагмент ДНК на участке хромосомы протяженностью 10 м.п.о. Современные методы гибридизации in situ с использованием метафазных хромосом, главным образом, метод FISH, локализуют полинуклеотидные маркеры в пределах 2–5 м.п.о. Более того, при гибридизации in situ с интерфазными хромосомами, в которых генетический материал находится в менее компактной форме, разрешающая способность хромосомных карт приближается к 100 т.п.о.
Точность хромосомных карт повышается и с использованием современных генетических методов. Например, способность ПЦР амплифицировать сегменты ДНК единичного сперматозоида позволяет исследовать большое число мейозов, как бы законсервированных в отдельных образцах спермы. В результате появляется возможность проверки взаимного расположения генетических маркеров, локализованных на хромосомных картах более грубыми методами.
Карты кДНК. Карты кДНК отражают положение экспрессирующихся участков ДНК (экзонов) относительно известных цитогенетических маркеров (бэндов) на метафазных хромосомах. Поскольку такие карты дают представление о локализации транскрибирующихся участков генома, в том числе и генов с неизвестными функциями, они могут быть использованы для поиска новых генов. Этот подход особенно полезен при поиске генов, повреждения которых вызывают заболевания человека, в том случае если приблизительная локализация таких участков хромосом уже предварительно проведена на генетических картах сцепления в результате семейного генетического анализа.
Физические карты высокого разрешения
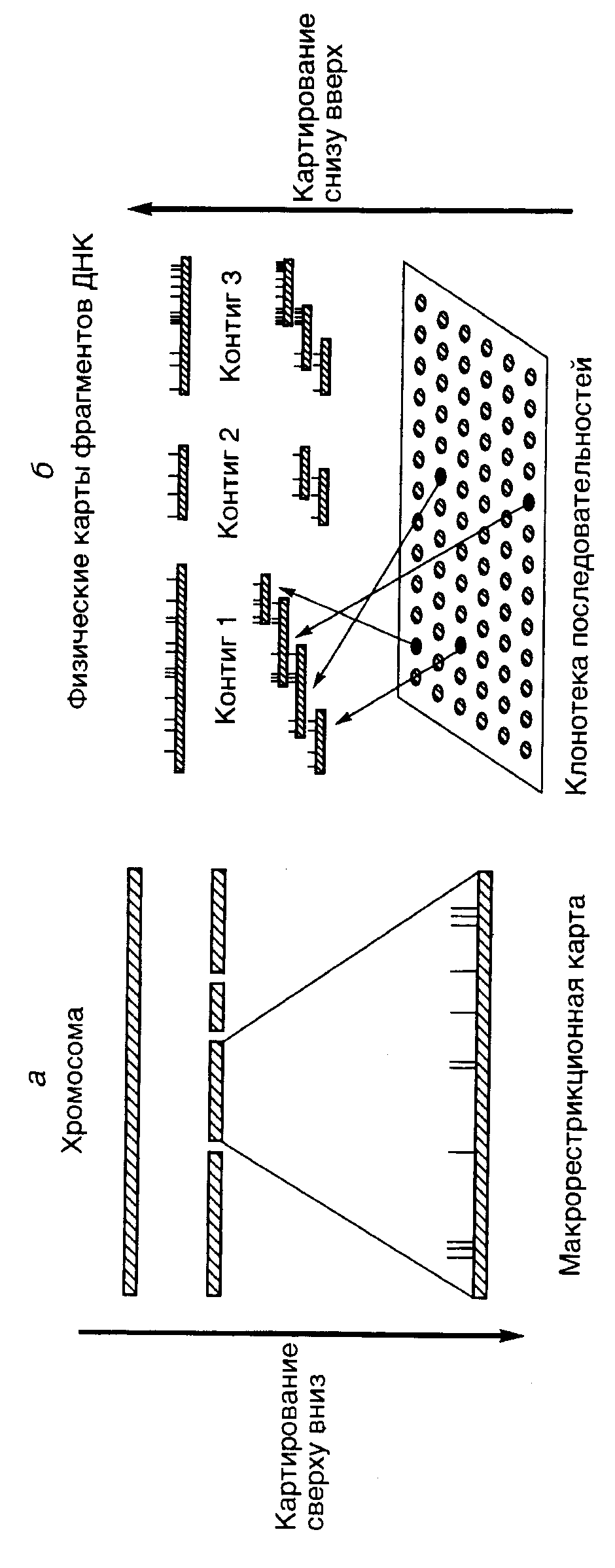
Рис. II.40. Две стратегии построения физических карт ДНК
а – стратегия "сверху вниз": ДНК целой хромосомы расщепляется крупнощепящими рестриктазами, для каждого из индивидуальных фрагментов ДНК строится рестрикционная карта; б – стратегия "снизу вверх", индивидуальные YAC-клоны после идентификации объединяются в контиги
В попытках построения карт генома человека высокого разрешения экспериментально реализуются два альтернативных подхода, получивших названия картирования сверху вниз (top-down mapping) и картирования снизу вверх (bottom-up mapping). При картировании сверху вниз (рис. II.40,а) исходным в анализе является препарат ДНК индивидуальной хромосомы человека. ДНК разрезается крупнощепящими рестриктазами (например NotI) на длинные фрагменты, которые после разделения электрофорезом в импульсном электрическом поле подвергаются дальнейшему рестрикционному анализу с другими рестриктазами. В результате получают макрорестрикционную карту, на которой достаточно полно представлены все последовательности исследуемой хромосомы или ее части, однако ее разрешение невысоко. На такой карте очень трудно локализовать индивидуальные гены. К тому же каждая индивидуальная карта редко охватывает протяженные сегменты ДНК (как правило, не более 1–10 м.п.о.).
При картировании генома человека снизу вверх (см. рис. II.40,б) на основе препарата суммарной ДНК генома или индивидуальной хромосомы получают серию случайных клонов протяженных последовательностей ДНК (10–1000 т.п.о), часть из которых перекрывается друг с другом. В качестве вектора для клонирования в этом случае часто используют искусственные минихромосомы бактерий (BAC) или дрожжей (YAC), подробно описанные в разделе 7.2.4. Серия частично перекрывающихся и дополняющих друг друга клонов образует непрерывную состыкованную (contiguous) последовательность нуклеотидов ДНК, получившую название контига (contig). Правильность полученных контигов подтверждают гибридизацией in situ (FISH) с одновременной их привязкой к определенным участкам исследуемых хромосом. Карты, основанные на контигах, представляют полную информацию о структуре отдельных сегментов хромосом и позволяют локализовать отдельные гены. Однако такие карты трудно применять для реконструкции целых хромосом или протяженных их участков из-за отсутствия соответствующих клонов в имеющихся клонотеках генов.
Основная проблема, которую приходится решать при использовании обоих подходов к построению физических карт высокого разрешения, – объединение разрозненных фрагментов ДНК в непрерывные последовательности нуклеотидов. Чаще всего для этого применяют специальные клонированные фрагменты ДНК, получившие название связующих (linking) клонов. Фрагменты ДНК из связующих клонов содержат в своих внутренних частях последовательности нуклеотидов крупнощепящих рестриктаз и, следовательно, представляют собой места стыковки фрагментов ДНК, используемых на первых этапах физического картирования. Гибридизацией по Саузерну, при проведении которой в качестве зондов используют фрагменты ДНК связующих клонов, определяют фрагменты ДНК физических карт, содержащие последовательности нуклеотидов окрестностей сайтов рестрикции крупнощепящих рестриктаз. Если два таких фрагмента найдены, то соответствующий связующий клон перекрывает оба этих фрагмента и является их частью. Связующие клоны, в свою очередь, отбирают из клонотек генов с помощью зондов, которые представляют собой последовательности нуклеотидов сайтов рестрикции крупнощепящих рестриктаз.