

Векторы для переноса днк в клетки животных и растений
Все основные принципы, используемые при конструировании бактериальных векторов, применимы и для получения векторов эукариотических клеток. Как и в случае бактерий, эукариотический вектор представляет собой небольшую молекулу ДНК, способную автономно реплицироваться в клетках животных. Помимо последовательностей нуклеотидов, обеспечивающих репликацию, эукариотические векторы могут содержать гены, используемые в качестве селектируемых маркеров, а также один или несколько уникальных сайтов рестрикции, по которым производится встраивание клонируемых последовательностей нуклеотидов ДНК. Поскольку непосредственное клонирование рекомбинантных ДНК в клетках животных было бы дорогостоящей и малоэффективной процедурой, эукариотические векторы используют, как правило, для получения экспрессии уже клонированных последовательностей нуклеотидов в клетках животных и растений, а сам процесс клонирования проводят в бактериях. Следовательно, эукариотические векторы, помимо всего прочего, должны быть челночными векторами. Для экспрессии в клетках рекомбинантные ДНК помещают под контроль регуляторных элементов, узнаваемых и используемых ферментативными системами эукариотических клеток.
В отличие от бактерий в клетках эукариот внехромосомные, автономно реплицирующиеся генетические элементы типа бактериальных плазмид встречаются редко. Поэтому при конструировании векторов, способных реплицироваться и осуществлять экспрессию в клетках животных, чаще всего используют регуляторные генетические элементы вирусов животных или, в зависимости от задач, эукариотических генов домашнего хозяйства, а также генов, для которых характерна тканеспецифическая экспрессия.
Экспрессирующий вектор pKSV-10. Структура теоретически возможного эукариотического вектора для экспрессии клонированных генов в клетках животных представлена на рис. II.12, из которого видно, что эукариотический вектор сохраняет все основные генетические элементы бактериальных векторов. Это, прежде всего, репликатор (область начала репликации ori), распознаваемый репликативными системами эукариотической клетки. В роли репликатора чаще всего используют соответствующие последовательности нуклеотидов вирусов животных (например вируса SV40 или вируса Эпштейна–Барр). Особенностью функционирования такого репликатора является потребность в вирусных белках для инициации репликации. Так, в случае SV40 – это T-антиген, тогда как у вируса Эпштейна-Барр – белок EBNA (Epstein-Barr nuclear antigen). Для того чтобы репликация векторов, в которых использованы вирусные репликаторы, осуществлялась без вируса-помощника, получили специальные линии клеток, которые стабильно продуцируют соответствующие вирусные белки. Например, в клетках линии COS синтезируется T-антиген вируса SV40, а клетки линии Hep-EBNA-2 экспрессируют ген EBNA.
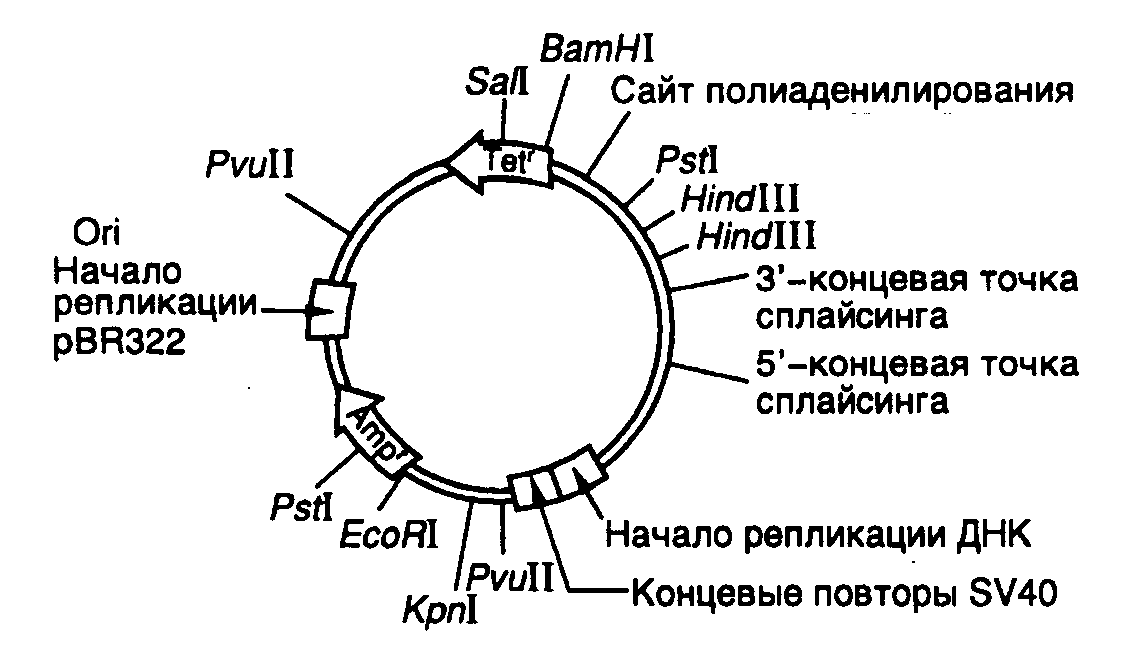
Рис. II.12. Эукариотический вектор pKSV-10
Часть бактериальной плазмиды, содержащей область начала репликации ori, соединена с транскрипционной единицей вируса SV40, 5’-концевая часть которой включает ранние промотор и репликатор вируса SV40, а 3’-конец содержит сайт полиаденилирования и интрон большого T-антигена. Последовательности чужеродной ДНК, которые необходимо экспрессировать в клетках животных, клонируют поBglII-сайту рестрикции. Кроме того, в векторе имеются уникальные сайты для рестриктазKpnI,BamHI,SalI иEcoRI. Вектор может реплицироваться в клетках E. coli и в клетках обезьян линии COS
В ряде случаев гены, необходимые для репликации эукариотического вектора, вводят непосредственно в вектор. Так же как и бактериальный, эукариотический вектор должен содержать один или несколько уникальных сайтов рестрикции для клонирования требуемой последовательности нуклеотидов. В экспрессирующем векторе полилинкер располагают непосредственно за генетическими элементами, регулирующими транскрипцию (наибольшее значение из них имеют промотор и энхансер), перед сайтом полиаденилирования и терминатором транскрипции. Структура других кассет генов, обеспечивающих экспрессию рекомбинантных генов в клетках животных, будет рассмотрена в разделе 7.4.6.
Векторы растений. Специфика векторов для экспрессии рекомбинантных генов в клетках растений обусловлена главным образом регуляторными последовательностями нуклеотидов, которые обеспечивают эффективную транскрипцию генов в растительных тканях. Например, в известном векторе pCaMVCAT используются промотор и сайт полиаденилирования генов вируса мозаики цветной капусты, под контролем которых находится ген бактериальной хлорамфениколацетилтрансферазы (cat). Этот ген-репортер применяется для оптимизации условий введения векторной ДНК в клетки растений и может быть заменен на любой другой ген. Вектор pCaMVCAT является челночным, т.е. может реплицироваться как в клетках E. coli, так и растений. В первом случае селектируемым маркером служит, как обычно, ген устойчивости к ампициллину.
Вектор pCaMVCAT принадлежит к большой группе так называемых DMGT-векторов (DNA-mediated gene transfer), осуществляющих опосредованный ДНК перенос генов в клетки растений. Это означает, что в данном случае перенос генов в клетки растений опосредован ДНК вектора, так как они находятся в составе векторной молекулы. Для получения стабильных линий растительных клеток, которые бы содержали рекомбинантные ДНК в интегрированном в хромосомы состоянии и активно экспрессировали его, часто в состав DMGT-векторов вводят дополнительные селектируемые маркеры, которые придают клеткам, включившим экзогенную ДНК, определенный, легко обнаруживаемый фенотип. Такими маркерами могут быть, например гены неомицинфосфотрансферазы II (NPT II), гигромицинфосфотрансферазы (HPT), дигидрофолатредуктазы, в результате экспрессии которых клетки приобретают устойчивость к канамицину, гигромицину B или метотрексату соответственно.