

Одомашнивание
Многие виды растений и животных сильно изменяются в процессе одомашнивания, при котором искусственный отбор сочетается с естественным. История выведения сортов культурных растений и пород домашних животных даёт множество хороших примеров эффективности селекции. Дарвин в «Происхождении видов» перечисляет различия между диким скалистым голубем и домашним голубем, в том числе такими специализированными породами, как турман и дутыш, в качестве примера глубоких эволюционных изменений, которые может вызвать искусственный отбор. Нетрудно вспомнить и другие примеры: собака, домашний скот, томаты, тыква и т. п.* В следующем разделе мы рассмотрим один из таких примеров — кукурузу.
Эволюция кукурузы
Кукуруза, или маис (Zea mays),— один из представителей трибы Maydeae семейства злаков (Gramineae) — резко отличается от всех других членов своей трибы и семейства по ряду признаков, в частности по расположению органов размножения и по строению колоса, или початка. Кукурузный початок — очень сложная и высокоспециализированная часть растения; ничего подобного нет ни у одного другого представителя семейства злаков. Но тем не менее современная кукуруза с её своеобразным початком и другими признаками происходит от какого-то дикого злака из тропических и субтропических областей Америки.
Соответствующие данные были получены в результате сравнительно-морфологических, таксономических, генетических, цитогенетических, этноботанических, археологических и палинологических исследований. В сборе этих данных участвовали многие ученые. Все они согласны с высказанным выше общим заключением, однако в том, что касается конкретной филогении, существуют различные точки зрения; к этому вопросу мы вернемся позднее*.
У Zea mays тычиночные и пестичные колоски находятся в разных соцветиях, колосьях и метелках соответственно. Кукуруза, таким образом, однодомное растение, что необычно для злаков. Несущая пыльцу метелка располагается на вершине кукурузного стебля, а початки, в которых находятся зерна, развиваются в пазухах листьев в средней части стебля (рис. 12.3). Початок кукурузы представляет собой колос, состоящий из толстого центрального стержня, несущего много рядов пестичных колосков, а позднее — столько же рядов зрелых зерен. Початок в целом покрыт чехлом из модифицированных листьев (листовые обвертки). Зерна прочно прикрепляющиеся к стержню, голые, т, е. не покрыты пленками (шелуха или мякина), как у других злаков (рис. 12.4, А, Б).
|
Рис. 12.3. Форма побега у двух сортов кукурузы (Zea mays) — зубовидной (А) и разрывной (Б). |

У других злаков каждое зерно покрыто отдельной пленкой и отделяется от соцветия, т. е. участвует в распространении семян в качестве самостоятельной единицы. Сочетание противоположных признаков у кукурузного початка — прочно прикрепленные голые зерна и листовые обвертки, одевающие початок в целом, — сильно снижает возможность распространения семян, однако эти же самые признаки очень ценны в сельскохозяйственном плане, поскольку они облегчают сбор урожая и сводят к минимуму потери зерна. Лучшие современные сорта кукурузы отличаются крупными размерами отдельных зерен и початка в целом (рис. 12.4, А). У некоторых примитивных сортов Zea mays, как и у её диких родичей, зерна мелкие, а початки маленькие (рис. 12.4, 5); при этом; зерна у них твердые и крем нистые, тогда как зерна лучших сортов мучнистые или сахаристые, так что их легче размолоть или разжевать.
Высокоурожайным новейшим сортам необходим мощный, хорошо облиственный стебель, чтобы синтезировать и запасать те вещества, ради которых их выращивают. Новые сорта отвечают этим требованиям. Современная кукуруза —гигант по сравнению с другими однолетними травянистыми растениями; она не ниже некоторых видов бамбука и гораздо выше примитивных сортов кукурузы (рис. 12.3). Эти и другие качества наиболее хорошо отселектированных сортов кукурузы полезны человеку, и он их создавал путем искусственного отбора на протяжении многих поколений.
|
Рис. 12.4. Початки трёх сортов кукурузы: зубовидной (А), аргентинской разрывной (Б) и пленчатой (В) (Mangelsdorf, 1958*). |

Разные земледельческие племена американских индейцев начали выращивать кукурузу уже по крайней мере 7000 лет назад (Mangelsdorf, 1974; 1983; Wilkes, 1972*). Центры культуры кукурузы возникли в восточной и юго-западной частях Северной Америки, в Мексике, Центральной Америке и в районах Анд. У сортов кукурузы, выращиваемых в этих центрах, всё ещё сохраняются некоторые примитивные признаки; о других признаках можно судить по остаткам давно исчезнувших сортов, найденным при археологических раскопках.
Разрывная (лопающаяся) кукуруза — относительно невысокое растение с несколькими побегами, отходящими от основания растения, и с небольшими початками, на которых сидят мелкие твердые зерна (рис. 12.3, Б и 12.4, Б). У пленчатой кукурузы проявляется другой примитивный признак: зерна покрыты пленками, как у других злаков (рис. 12.4, В).
|
Рис. 12.5. Продольный разрез нижней части ископаемого кукурузного початка из Бат-Кейв; верхний конец початка не сохранился. (Mangelsdorf, 1958.*) |
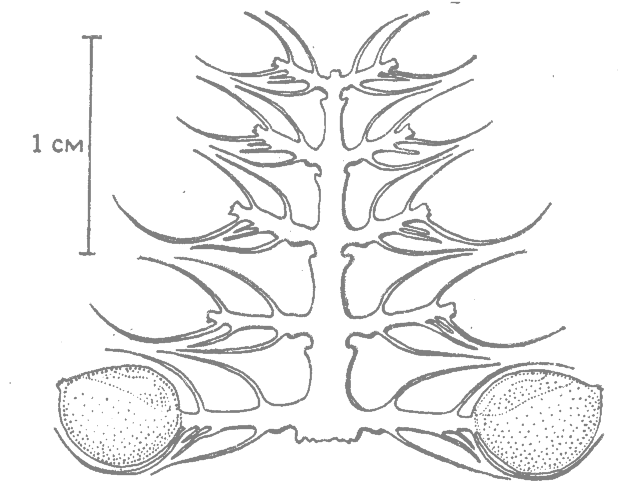
Важную находку представляет собой ископаемый кукурузный початок, найденный в Бат-Кэйв (шт. Нью-Мексико) — пещере, где жили первобытные земледельцы, выращивавшие примитивный сорт кукурузы. Этот початок маленький, меньше, чем у разрывной кукурузы. Зерна такие же мелкие, как у разрывной кукурузы, сидят на длинных плодоножках, отходящих от сравнительно тонкого центрального стержня, и покрыты пленками, как у пленчатой кукурузы (рис. 12.5). Возможно, что верхний конец колоса нес тычиночные колоски. При помощи радиоуглеродного и других методов было установлено, что возраст самых старых экземпляров кукурузы, найденных в Бат-Кэйв, составляет 3000— 4000 лет (Mangelsdorf, Smith, 1949; Mangelsdorf, 1958; 1974*). У более древних ископаемых початков, найденных в Теуакан- Воллей (Мексика), возраст которых достигает почти 7000 лет, также обнаружено сочетание признаков разрывной и пленчатой кукурузы; колосья были обоеполыми с тычиночными соцветиями на конце (Mangelsdorf et al., 1964; Mangelsdorf, 1974; 1983; Wilkes, 1978*).
Среди ещё более древних ископаемых остатков из Мехико найдены пыльцевые зёрна, сходные с пыльцевыми зёрнами современной кукурузы. Хотя определение на основании одних только пыльцевых зерен без других частей растения нельзя считать вполне надёжным, весьма вероятно, что найденная в Мехико пыльца принадлежит кукурузе, а не теосинте или какому-нибудь другому злаку. Она, возможно, представляет предковую форму кукурузы. Возраст этих пыльцевых зерен, согласно оценкам, равен 25000 — 80000 лет (Mangelsdorf, 1974; Mangelsdorf et al., 1978*).
Мангельсдорф (Mangelsdorf, 1958*) скрещивал современную разрывную кукурузу с пленчатой и получил синтетический сорт, в котором сочетались примитивные признаки обеих родительских форм. У этой синтетической пленчато-разрывной кукурузы узкое обоеполое соцветие, типичное для злаков, с женскими цветками в нижней и мужскими в верхней частях. Зерна покрыты пленчатыми чешуйками и легко отделяются от соцветия после созревания. Таким образом, синтетическая пленчато-разрывная кукуруза обладает соцветием, типичным для злаков (рис. 12.6,Б); она, вероятно, соответствует одной из возможных ранних стадий в эволюции кукурузы.
В тропических и субтропических областях Америки ближайшие родичи Zea mays представлены трёмя группами: 1) небольшой дикорастущий род Tripsacum (рис. 12.6 А); 2) сорняк Zea mexieana, или однолетняя теосинте (рис. 12.6 В); 3) недавно обнаруженные дикие многолетние теосинте, Zea perennis и Z. diploperennis. Все они считаются предками Zea mays.
Разные виды Tripsacum, в том числе Т. dactyloides, широко распространены в теплых областях Нового Света. В отличие от Zea mays и Z. mexieana, Tripsacum — многолетник и несёт обоеполые соцветия (рис. 12.6, А). Соцветие Tripsacum мало отличается от соцветия синтетической пленчато-разрывной кукурузы. Tripsacum может быть успешно скрещён с Zea mays, но не с Z. mexieana, и гибриды F1, если их удается получить, более или менее стерильны.
|
Рис. 12.6. Колосья трёх родичей современной кукурузы, обладающих примитивными признаками. A. Tripsacum dactyloides верхние колосья мужские, а нижние — женские. Б. Синтетическая пленчато-разрывная кукуруза — верхние колосья мужские, а нижние — женские. В. Zea mexicana — ветвящиеся ряды женских колосков (Mangelsdorf, 1958; Wilkes, 1967.*) |

Теосинте (Z. mexieana) — однолетнее растение, широко распространенное в Мексике и Центральной Америке, где оно часто встречается в качестве сорняка на кукурузных полях или вблизи них. Z. mays и Z. mexieana можно искусственно скрещивать, а в Мексике они скрещиваются в природе, причём гибриды F1 плодовиты. У Z. mexicana, так же как у Z. mays имеются от дельные тычиночные и пестичные соцветия, однако пестичные колоски имеют у нее вид разветвленной системы тонких колосьев, окруженных листовой обверткой. У Z. mexicana каждый из собранных в колос пестичных колосков защищён особой отдельной обверткой (плюской); у Z. mays такие плюски имеются, но они редуцированы и служат скорее для повышения прочности, чем для защиты. У Z. mexicana стержень колоса расчленяется, так что зерна легко отделяются друг от друга. У теосинте нет початка (рис. 12.6, б).
Большой интерес представляют две дикие многолетние формы теосинте, недавно обнаруженные в Халиско (Мексика), поскольку в группах травянистых растений, содержащих как однолетние, так и многолетние формы, многолетники обычно считают более примитивными. Одна многолетняя форма, тетраплоид, Z. perennis, была известна и раньше, но считалась вымершей, так что в сущности она была открыта вторично. Другая, диплоид Z. diploperennis, оставалась неизвестной науке до того, как была обнаружена в 1978 г. (Iltis et al., 1979; Doebley, Iltis, 1980*).
В общем считается, что предковые признаки кукурузы можно обнаружить у её родичей: Tripsacum, однолетней теосинте и многолетней Zea. Однако при попытках построить определённую филогению возникают затруднения и противоречия. Одна приемлемая филогенетическая гипотеза состоит в том, что род Zea происходит от Tripsacum (Doebley, Iltis, 1980*). Согласно второй гипотезе, Zea diploperennis при гибридизации с дикой кукурузой даёт теосинте (Mangelsdorf et al., 1978; Wilkes, 1982; Mangelsdorf, 1983*).
Что касается предк