

Синтез липидов из углеводов
Если количество углеводов, поглощенных с пищей за один прием, больше того, чем может быть запасено в виде гликогена, то избыток углеводов превращается в жиры. Начальная последовательность реакций совпадает при этом с обычным окислительным путем, т.е. сначала из глюкозы образуется ацетил-КоА, который используется в цитоплазме клетки для синтеза длинноцепочечных жирных кислот. Затем жирные кислоты запасаются в виде нейтральных жиров (триглицеридов), которые откладываются в разных частях тела. Когда требуется энергия, нейтральные жиры подвергаются гидролизу и жирные кислоты поступают в кровь, адсорбируются молекулами плазменных белков (альбуминов и глобулинов) и затем поглощаются различными клетками.
Циклы Кори и аланина
В клетках, не содержащих митохондрий (например, в эритроцитах), или в тканях при недостаточном снабжении кислородом (например, в активно работающих мышцах) АТФ (АТР) синтезируется за счет процесса превращений глюкозы в лактат (анаэробного гликолиза).
Лактат переносится кровью в печень, где в процессе глюконеогенеза с затратой АТФ вновь конвертируется в глюкозу (цикл Кори).
При интенсивной работе мышц максимально активируется гликолиз. Продукт гликолиза, пировиноградная кислота (пируват) накапливается в цитоплазме и недостаточно быстро поступает в митохондрии, если они из-за недостатка кислорода не готовы к окислению пирувата. В анаэробных условиях пируват в реакции, катализируемой лактатдегидрогеназой (заключительный этап гликолиза) восстанавливается до лактата. Одновременно НАДН (NADH), кофермент лактатдегидрогеназы, окисляется до НАД+ (NAD+), который вновь используется на окислительном этапе гликолитического пути. Этой реакции способствует относительно высокое отношение НАДН/НАД+ в мышечной ткани. Лактат диффундирует в кровь и поступает в печень, где конвертируется в глюкозу. Таким образом, образование лактата временно заменяет аэробный метаболизм глюкозы и частично переносит этот процесс из мышц в печень.
Обратная связь, подобная циклу Кори, существует в глюкозо-аланиновом цикле, в котором также участвует пируват. Глюкозо-аланиновый цикл берет начало с протеолиза белков. Образующиеся аминокислоты в результате трансаминирования в присутствии ферментов превращаются в α-кетокислоты, которые в основном включаются в цикл трикарбоновых кислот (цитратный цикл). Одновременно в реакции, катализируемой аланинтрансаминазой, аминогруппы из разных аминокислот переносятся на имеющийся субстрат, пируват. Образующийся аланин поступает в кровь и переносится в печень. Таким образом, цикл аланина служит каналом передачи азота и предшественников глюкозы в печень, которая является местом синтеза конечных продуктов азотистого обмена, например мочевины.
Роль инсулина в регуляции обмена веществ
Инсулин участвует в регуляции всех видов обмена веществ. Это основной гормон, обладающий гипогликемическим действием (снижению сахара крови также способствует соматостатин).
Он является полипептидным гормоном, образующимся в β-клетках поджелудочной железы. Главным стимулятором синтеза и секреции инсулина является глюкоза. Образующийся инсулин поступает в сосудистое русло, где он частично остается в свободном виде, а частично образует комплексы с белками крови.
Инсулин опосредует поступление и метаболизм глюкозы в ткани, имеющие рецепторы к инсулину (инсулинзависимые ткани), рис. 7.
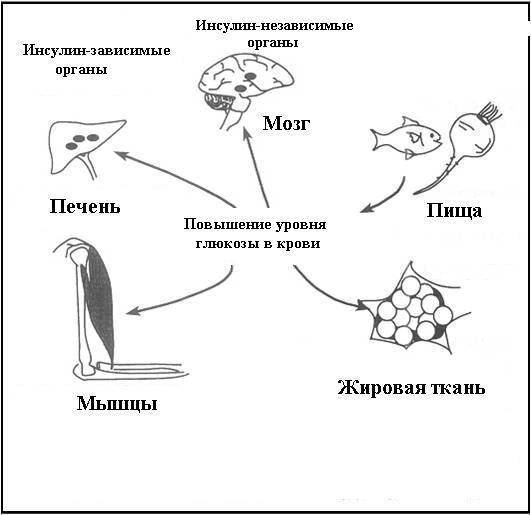
Рис. 7. Инсулин-зависимые и инсулин-независимые органы.
К инсулинзависимым тканям относятся мышечная, жировая ткань, печень и островковый аппарат поджелудочной железы. В эти ткани глюкоза поступает путем пассивного переноса или облегченной диффузии. Остальные ткани являются инсулиннезависимыми и поступление глюкозы в них происходит без участия инсулина по градиенту концентрации.
Активируя гексокиназу, в клетках инсулинзависимых тканей инсулин обеспечивает фосфорилирование глюкозы, превращение ее в Гл-6-фосфат и дальнейшее превращение. Инсулин стимулирует гликолиз, цикл Кребса, пентозофосфатный путь обмена глюкозы и угнетает глюконеогенез. Инсулин усиливает гликогенез в результате активации гликоген-синтетазы и тормозит гликогенолиз.
Кроме глюкозы инсулин также опосредует поступление в клетки аминокислот и электролитов. Он активирует синтез белка, жира и тормозит их распад, препятствует избыточному холестерол- и кетообразованию. Инсулин также участвует в регуляции водно-электролитного обмена (обладает водо- и солесберегающим действием), участвует в регуляции кислотно-основного состояния (препятствует развитию ацидоза).
Суточная потребность в инсулине – 40 единиц, а его содержание в поджелудочной железе здорового человека составляет 150-250 Ед. Инактивация инсулина происходит преимущественно в печени и почках под влиянием инсулиназы.
По влиянию на уровень глюкозы другие гормоны (глюкагон, соматотропный гормон (СТГ), пролактин, адреналин и норадреналин, глюкокортикоиды) относятся к группе контринсулярных гормонов. Глюкагон синтезируется в α-клетках поджелудочной железы. Механизм гипергиликемического действия глюкагона связан с усилением гликогенолиза в печени. Аналогичным действием обладают СТГ, пролактин, адреналин и норадреналин. Гипергликемическое действие глюкокортикоидов преимущественно связано с их стимулирующим влиянием на глюконеогенез.