

«Феномен просветления плазмы крови»
После принятия жирной пищи плазма выглядит мутной (хилезной). Через 12 часов она просветляется. В капиллярах легких, жировой, мышечной ткани и селезенке под влиянием фермента эндотелиальной липопротеинлипазы (ЛПЛ, «фактор просветления») происходит расщепление триглицеридов, входящих в состав хиломикронов, до свободных жирных кислот и глицерина – «феномен просветления плазмы крови».
В активации ЛПЛ принимает участие гепарин из тучных клеток, расположенных по ходу сосудов, а также апопротеин апо-С-II, входящий в состав хиломикронов. Дефицит апо- С-II является причиной одного из видов гиперлипопротеинемий – хиломикронемии (см. ниже). Феномен просветления осуществляется также в жировой ткани и других органах.
Нерасщепленные хиломикроны (ремнанты хиломикронов) продолжают циркуляцию и захватываются печенью, где происходит их окончательное расщепление под влиянием печеночной липопротеинлипазы. Также в соединении с альбуминами в печень поступают неэтерифицированные жирные кислоты, образовавшиеся в результате распада триглицеридов хиломикронов в сосудистом русле.
Патология межуточного обмена липидов. Роль печени
Одним из этапов липидного обмена в печени является образование липидов (эндогенных триглицеридов, фосфолипидов, холестерола, кетоновых тел). За исключением витамина D, все встречающиеся в организме животных стероиды (производные сложных спиртов) – холестерол (холестерол), желчные кислоты, мужские и женские половые гормоны и гормоны надпочечников синтезируются в организме. Исходным материалом для синтеза служит ацетил-КоА.
При связывании эндогенных триглицеридов аполипопротеинами образуются липопротеиновые комплексы очень низкой плотности – ЛОНП. Они попадают в кровь и переносятся в другие ткани, прежде всего в жировую и мышечную ткань (рис.11).
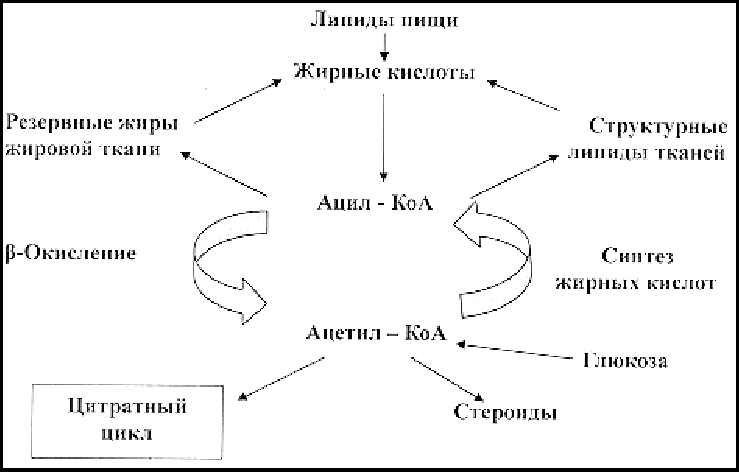
Рис.11. Межуточный обмен липидов.
Окисление жирных кислот протекает путем последовательного отщепления от молекулы жирной кислоты двууглеродного фрагмента с образованием ацетилкофермента A (ацетил-КоА) и одновременной передачей двух пар электронов в цепь переноса электронов. Образовавшийся ацетил-КоА – компонент цикла трикарбоновых кислот, судьба которого в дальнейшем не отличается от судьбы ацетил-КоА, поставляемого углеводным обменом. Таким образом, механизмы синтеза АТФ при окислении, как жирных кислот, так и метаболитов глюкозы практически одинаковы.
Если организм животного получает энергию почти целиком за счет одного только окисления жирных кислот (голодание, сахарный диабет), то скорость образования ацетил-КоА превышает скорость его окисления в цикле трикарбоновых кислот. В этом случае лишние молекулы ацетил-КоА реагируют друг с другом, в результате чего образуются в конечном счете ацетоуксусная и β-оксимасляная кислоты.
Кетоновые тела (содержание в норме 0,08-0,43 mМ) поступают из печени в кровь. Во второй период голодания наряду с жирными кислотами 3-гидроксибутират и ацетоацетат в являются основными энергоносителями. Ацетон, не имеющий метаболической ценности, удаляется через легкие. Если биосинтез кетоновых тел превышает потребности организма, они накапливаются в крови (кетонемия) и выводятся с мочой (кетонурия). Возрастание концентрации кетоновых тел вызывает изменение рН в крови (кетоацидоз). Кетонурия и кетоацидоз могут быстро привести к электролитному сдвигу (нарушению ионного гомеостаза), потере сознания вследствие развития кетоацидотической комы и опасны для жизни.
При патологии печени происходят изменения межуточного обмена липидов, проявляющие уменьшением β-окисления жирных кислот, усилением кетообразования, синтеза холестерола, уменьшением образования фосфолипидов.
Уровень кетоновых тел достигает при длительной мышечной работе не >0,1mМ, голодании – до 6,45 mМ, сахарном диабете – до 40-50 mМ.