

7.2.2 Ценобластическая организация
Некоторые водоросли и грибы имеют много ядер, но не разделены на клетки. В их вегетативных телах, чаще всего имеющих трубчатую форму, иногда довольно крупных и высокоорганизованных (рис. 7.4), каждое ядро образует вместе со своей «сферой влияния» в протоплазме энергиду.
Плазмодии – многоядерные массы протоплазмы, образующиеся в результате слияния голых клеток (настоящие плазмодии у слизистых грибов - миксомицетов) или просто их группировки (псевдоплазмодии у миксобактерий). Они могут как целое медленно перетекать или переползать с места на место.
7.2.3 Объединения клеток
Объединения клеток представляют собой ступени на пути к многоклеточности.
Ценобии – комплексы слабо объединенных клеток – возникают в результате клеточного деления. Клетки остаются связанными студенистым материалом, набухшими клеточным стенками или целлюлозными домиками (рис. 7.4), но не образуют функционального единства и всегда могут разойтись. Ценобии встречаются уже у прокариот (рис. 7.4).
Агрегационные объединения (рис. 7.4) образуются в результате временного соединения одноклеточных организмов. Они могут как угодно распадаться на более мелкие единицы.
Колонии возникают путем клеточного деления. Клетки в них образуют морфологическое и функциональное единство (рис. 7.4). Их цитоплазмы соединены между собой и могут реагировать координирование (синхронное биение жгутиков).
В высокоорганизованных колониях, например в шаровидной водоросли Volvox (рис. 7.4), имеет место разделение функций (есть вегетативные клетки, обеспечивающие движение, и генеративные, служащие для размножения), и их можно уже считать первыми многоклеточными организмами.
7.2.4 Многоклеточные организмы без истинных тканей
Все многоклеточные растения можно объединить под названием Metaphyta (сюда входят Thallophyta, Bryophyta и Cormophyta), а животных – под названием Metazoa (Porifera и Histozoa). Bryophyta, Cormophyta и Histozoa обладают настоящими тканями.
Таллофиты (Thallophyta). Таллом – это многоклеточное (или ценобластическое) вегетативное тело, не подразделенное на корень, стебель и листья. Таллофиты – растения с талломом – не составляют единой таксономической группы, а входят в состав различных групп водорослей и грибов (рис. 7.5).
|
|
|
|
1 |
2 |
3 |
4 |
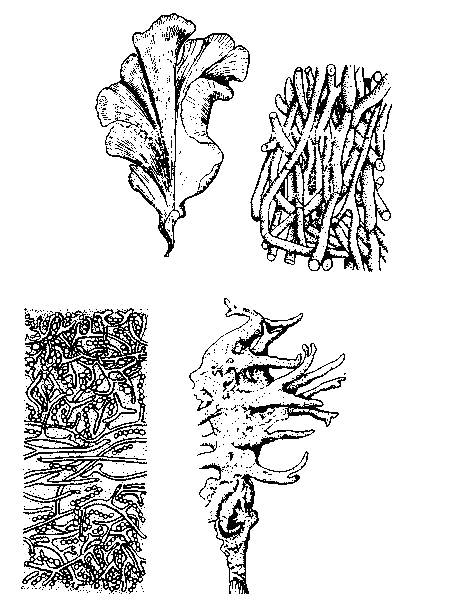

Рис. 7.5. Таллофиты и губки. 1 – зеленая водоросль Ulva (х1/3);
2 – плектенхима в ножке плодового тела белого гриба, Boletus edulis (х200);
3 – поперечный срез лишайника Collema (X200); 4 – пресноводная губка Spongilla lacustris (XI/2)
В нитчатых талломах клетки, размножаясь поперечным делением, образуют один ряд. Плоские талломы возникают из нитчатых в результате продольного деления клеток (рис. 7.5). У более высокоразвитых форм появляется полярная дифференцировка: у них есть растущий конец таллома и прикрепляющийся к субстрату базальный ризоид (рис. 7.5). Другой шаг вперед – ветвление таллома. Таллом грибов – мицелий – состоит чаще всего из множества обильно ветвящихся нитей – гиф.
Талломные нити могут объединяться в ложные ткани (у красных водорослей, в плодовых телах высших грибов): переплетаясь, они образуют плектенхиму (плетеную ткань), а срастаясь – псевдопаренхиму (рис. 7.5).
Лишайники – это симбиотические автотрофные объединения водоросли с грибом. Обычно в плектенхиму гриба внедрены одноклеточные синезеленые или зеленые водоросли (рис. 7.5).
Губки (Porifera) помимо клеток, служащих для размножения, обладают различными типами соматических клеток, которые, однако, не объединяются в органы или настоящие ткани (рис. 7.5). Только на поверхности тела клетки могут образовывать подобие эпителия. Примечательна способность уже дифференцировавшихся клеток к превращению. Они скреплены лишь рыхло и легко могут снова выйти из объединения. Большинство клеток способно к амебоидному движению. Губки – прикрепленные (сидячие), в основном морские организмы.