

11.2.2 Основы популяционной генетики
Изменившиеся организмы будут иметь значение для эволюции только в том случае, если их станет достаточно много и они смогут образовать новые размножающиеся сообщества (популяции), виды и более высокие таксономические единицы. Изменения популяции зависят от разных факторов, в том числе от способа размножения. Различают аутогамные и аллогамные популяции (в которых размножение происходит соответственно путем самооплодотворения и перекрестного оплодотворения). Аутогамные популяции свойственны многим растениям (например, пшенице и ржи). В них преобладают гомозиготные организмы. Гетерозиготные особи получаются лишь при скрещивании генетически различных гомозигот или после мутаций. В ряду поколений доля гомозигот растет, так как они дают только гомозиготное потомство, а в потомстве гетерозигот опять-таки выщепляются гомозиготы.
В аутогамной популяции среди гомозигот (доля которых возрастает в результате самооплодотворения) и немногочисленных гетерозигот различные типы, возникшие в результате мутаций или скрещивания, встречаются с разной частотой. Их доля зависит от частоты скрещиваний, частоты появления мутантов, условий отбора и других факторов, а также от времени появления нового аллеля. Каждая популяция изменяет свой состав, если только условия внешней среды и отношения внутри популяции не привели к равновесию.
Аллогамные популяции свойственны почти всем животным и многим растениям. Состав аллелей в них определяется не только мутациями, но и в очень значительной степени рекомбинациями. Так как потомство образуется в результате скрещиваний, доля гетерозигот относительно велика. Состав популяции при этом подчиняется закону, выведенному Харди и независимо от него Вейнбергом.
Если в популяции, состоящей из типов а+а+, а+а- и а-а-, обозначим через р долю аллеля а+ и через q долю а-, то для популяции будет характерно определенное соотношение р : q. В идеальных популяциях соотношение типов гамет р : q одинаково у обоих полов. Поэтому скрещивания дают расщепление по формуле p2--2pq--q2:
-
p
q
p
p2
pq
q
qp
q2
Здесь р2 – доля особей а+а+, 2р – доля особей а+а- и q2 – доля особей а-а-. Соотношение аллелей р : q и соотношение типов р2 : 2pq : q2 в идеальной аллогамной популяции остаются постоянными.
Эти закономерности, а также постоянство соотношения типов верны только для идеальных популяций, в которых число особей очень велико, существует панмиксия (равные шансы скрещивания для всех особей), нет отбора, нет мутаций, нет миграции (эмиграции и иммиграции особей) и нет других случайно изменяющихся факторов. Поскольку на естественные популяции воздействует множество переменных факторов, частота разных аллелей, как правило, в ряду поколений изменяется.
11.2.3 Возникновение наследственных вариантов
Особи с наследственными изменениями возникают в результате мутаций и скрещиваний (рекомбинации) организмов с разными аллелями или генами. Пока особенности размножения, морфологии или экологии изменены лишь незначительно, новые популяции остаются в составе исходного вида. Только после явного возникновения механизмов изоляции, отделяющих их от исходного вида, можно говорить о новом виде (или более высоком таксоне).
В результате генных мутаций возникают новые аллели, которые приводят к неблагоприятным, благоприятным или безразличным изменениям исходного типа. Мутанты с неблагоприятными изменениями более или менее быстро элиминируются, если это гаплоидные организмы или диплоиды с доминантной мутацией. При рецессивной мутации новый аллель может сохраниться в популяции в гетерозиготном состоянии. Аллель, который в гомозиготном состоянии вызывает неблагоприятный эффект (как, например, ген серповидноклеточной анемии), в гетерозиготном состоянии иногда может создавать селективное преимущество (в данном примере он снижает восприимчивость к малярии). У нейтральных мутантов наследственное изменение вначале не проявляется в фенотипе. Основываясь на измененных таки образом аминокислотных последовательностях, удавалось строить эволюционные схемы и делать выводы о родственных связях между организмами.
Филогенетическое дерево, представленное на рис. 11.6, основано на данных об изменениях в гене rРНК 16S (малой субчастицы рибосом). Оно свидетельствует о происхождении прокариот и эукариот от общих предков.
Степень сходства выражали как отношение
![]() ,
,
где А и В – сравниваемые виды. Величине SAb соответствует на схеме положение самой нижней горизонтальной линии, соединяющей оба вида. При SAb = 1,0 совпадение было бы полным.
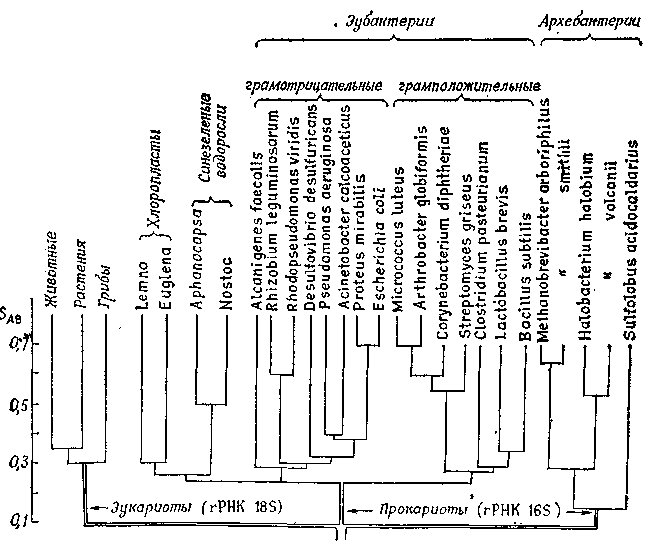
Pиc. 11.6. Филогенетическое дерево, построенное по данным о сходстве олигонуклеотидов, получаемых из rРНК малой субъединицы рибосом. (По Fox et al., с изменениями).
О значении благоприятных мутаций часто можно судить по сравнительно быстрому распространению их носителей, особенно в том случае, если они хорошо приспособлены к изменившимся условиям. У березовой пяденицы (Biston betularia) кроме светлой нормальной формы встречается темный мутант (саrbonaria). Эти две формы различаются аллелями одного гена и при скрещивании дают менделевское расщепление. В Англии в областях с сильным задымлением атмосферы преобладает мутантная форма, а в районах с более чистым воздухом – нормальная, т.е. в каждом случае та, которая менее заметна для врагов.
Одна или несколько генных мутаций, как правило, приводят лишь к внутривидовой эволюции. Границы вида преодолеваются только в результате многочисленных (или особенно важных) мутаций, если возникающая при этом новая форма изолируется от исходного вида репродуктивными барьерами.
Хромосомные мутанты при скрещивании с исходной формой дают иногда такое же менделевское расщепление и имеют в этом случае такое же значение для эволюции, как генные мутанты.
Для возникновения новых функций большое значение имеют мутации в дуплицированных участках. Удвоенные гены или части генов кодируют вначале совершенно одинаковые пептиды или части пептидов. Новая информация возникает тогда, когда в дуплицированных участках независимо друг от друга происходят мутации. Примером может служить возникновение изоферментов (ферментов с очень сходной функцией). Почти неизмененные дупликации могут увеличивать продуктивность важных генов; например, гены рибосомальной РНК.
В процессе эволюции происходили крупные изменения числа хромосом. У современных организмов на один геном приходится от 1 до 500 и более хромосом. Их число часто бывает полиплоидным или анэуплоидным (рис. 11.7).
Полиплоиды составляют 30-50% всех цветковых растений (это в большинстве случаев аллополиплоиды). После полиплоидизации в результате неправильного расхождения хромосом может произойти анэуплоидизация. Анэуплоиды могут возникать и из триплоидов.
В ходе эволюции произошел огромный рост количества ДНК в геноме: у бактерий один геном содержит 106-107 пар нуклеотидов, у млекопитающих – около 5-109. Однако у высших организмов около 30% ДНК повторяется, у бактерий же повторяющейся ДНК всего лишь около 0,3%.
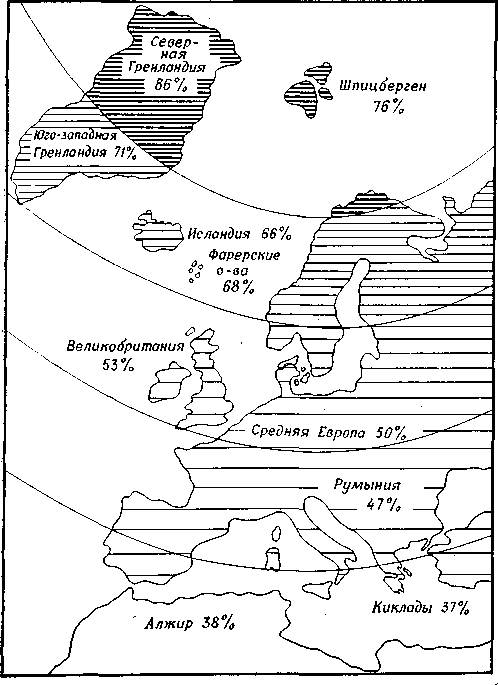
Рис. 11.7. Доля полиплоидных растений во всей растительности. (Vogel)
Скрещивания – гораздо более частые события, чем мутации, особенно у видов с перекрестным оплодотворением. Приводя к рекомбинации генов, они значительно повышают многообразие форм внутри вида. Число возможных новых комбинаций обычно так велико, что они даже не могут реализоваться при имеющейся численности особей данного вида. В поколении F2 при полностью фертильных скрещиваниях и наличии п разных аллелей оно составляет 2n-2, т.е. при 5 разных аллелях уже равно 30. Среди них могут быть и такие типы, которые морфологически и физиологически настолько отклоняются от исходных форм, что при изменении экологических условий могут дать начало новым видам.