

12.3 Организм и среда
Каждый организм обладает способностью реагировать в соответствии со своей генетической конституцией на окружающую среду, использовать ее факторы для своего существования и развития или, по крайней мере, переносить их воздействие. Эта экологическая потенция определяется наследственной нормой реакции по отношению к каждому фактору среды. Норма реакции у каждого вида характеризуется определенным положением и диапазоном (широтой) на шкале интенсивности данного фактора – экологической валентности. Экологическая потенция охватывает диапазон от нижнего пессимального предела (минимума) до верхнего пессимального предела (максимума) с оптимумом где-то посередине (рис. 12.7). В пессимумах способность реагировать снижается до нуля, за ними наступает абсолютное торможение процесса (реакции), часто даже необратимое повреждение организма. Широта экологической потенции (от минимума до максимума) у эврипотентных видов велика, у стенопотентных мала. Смотря по тому, где лежит оптимум для стенопотентного вида — в нижней, средней или верхней части всего диапазона, — различают олиго-, мезо- и полипотентные виды. Положение и широта нормы реакции зависят от возраста, пола и фазы развития и различны для разных процессов (активности, метаболизма, развития и т.д.). Границы экологической потенции могут (обычно лишь ненамного) сдвигаться в результате модификации, т.е. индивидуального приспособления (привыкания) или мутации, т.е. генетического приспособления
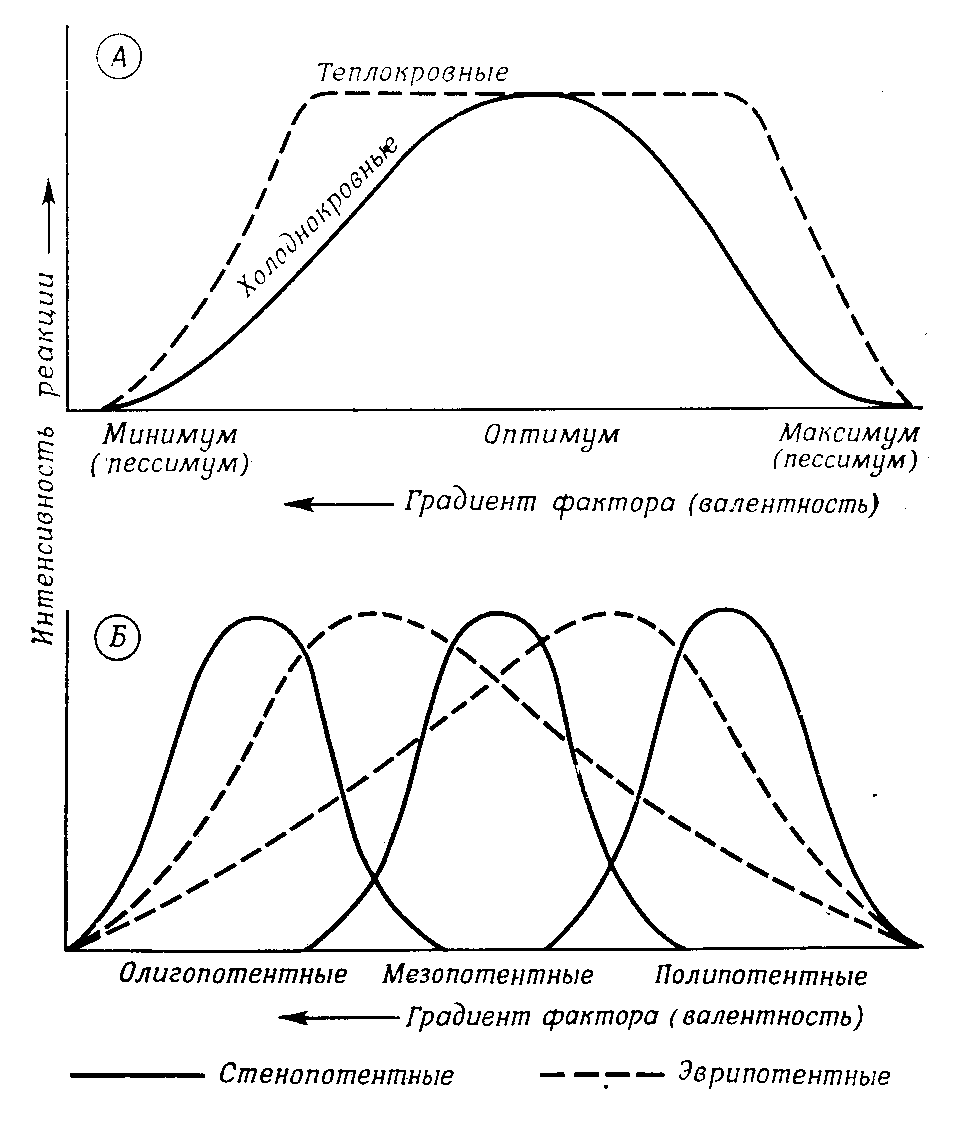
Рис. 12.7. Экологическая потенция в сопоставлении с областью валентности фактора среды А. Общая схема для пойкилотермных и гомойотермных животных. Б. Различные положения и ширина области потенции (норма реакции)
12.3.1 Фактор температуры
Температура влияет на энергетику всех жизненных процессов. Так как в основе всех реакций живого организма, зависящих от температуры, в конечном счете лежат биохимические процессы, для них в основном верно правило зависимости скорости реакции от температуры – закон Вант-Гоффа, согласно которому при повышении температуры на 10°С реакция ускоряется в 2-3 раза. Однако под влиянием эндогенной нормы реакции и лимитирующих внешних факторов соответствующая экспоненциальная кривая рано или поздно переходит в кривую с оптимумом.
Активность животных ограничивается пессимумами, при которых наступает обратимое тепловое или холодовое окоченение. У насекомых повышение температуры вызывает вначале медленные, некоординированные движения, в физиологической области (оптимум) приводит к полностью управляемой активности, а при дальнейшем повышении – к чрезмерно быстрым, некоординированным, суматошным движениям.
Главная точка приложения для воздействия температуры – обмен веществ. Для ассимиляции, как и для диссимиляции, существуют специфические температурные пределы и специфический оптимум.
Эвритермные организмы (с широкой экологической потенцией) – это, например, актиния Actinia equina, которая может жить как в тропических, так и в арктических морях; карпы, способные жить и в холодных, и в теплых пресных водоемах. Стенотермные виды приспособлены к узкому диапазону температур. Среди них политермны (т.е. приспособлены к теплу) почти все рифообразующие кораллы, термиты, тараканы и другие животные; олиготермны (приспособлены к холоду) гренландский кит, самцы зимней пяденицы, ручьевая форель, сардиновые рыбы.
Подвижные организмы могут использовать внешние источники тепла или избегать их, изменяя позу по отношению к источнику теплового излучения (гелиорегуляция) или отыскивая места, более подходящие в отношении тепла (термотаксис). Холодным утром кузнечики подставляют бока солнечному свету, а дневные бабочки расправляют крылья; в полуденную жару они, сложив крылья, располагаются параллельно лучам. Высокие компасные термитники, так же как листья компасных растений, ориентированы широкими сторонами на восток и запад (что дает выигрыш в инсоляции утром и вечером), а узкими – на север и юг (так что нет перегрева в середине дня).
Термопреферендум (предпочтительная температура) специфичен для каждого вида; это та температура, при которой не расходуется энергия ни для производства тепла, ни для его отдачи. Организм обычно находит такие условия путем фоботаксиса, в результате беспорядочных передвижений (методом проб и ошибок).
Предпочитаемая температура (ПТ), в общем, соответствует экоклимату нормального местообитания данного вида. У олиготермной форели (из верховий текучих вод) она составляет 10,5°С, у эвритермных карпов (стоячие воды) – 21,3. Она может быть различной на разных стадиях развития. Строительство гнезд служит у наземных беспозвоночных, птиц и млекопитающих прежде всего для сохранения тепла, выработанного в организме или полученного от внешнего источника. Общественные пчелы летом поддерживают в улье удивительно постоянную температуру – от 34,5 до 35,5°С, согревая его движением крыльев или мышц либо охлаждая вентиляцией и испарением принесенной воды.
Только гомойотермные (теплокровные) животные-птицы и млекопитающие – сравнительно независимы от окружающей температуры и создают свой собственный «внутренний климат», поддерживая постоянную температуру тела (37-39°С у млекопитающих, 41-42°С у птиц). Поэтому кривые их реакции на температуру образуют почти ровное плато между пессимумами (рис. 12.7). И все же у этих животных тоже есть предпочитаемые температуры (такие, при которых им приходится тратить минимум энергии на терморегуляцию); они обладают такими приспособлениями, как сезонные изменения густоты и длины шерсти, а для выращивания молоди используют теплоизолирующие материалы. Поэтому многие из них могут жить почти во всех климатических зонах (например, пума и тигр – от субарктики до тропиков).
При сравнении родственных видов из теплых и холодных областей часто можно отметить увеличение размеров тела (правило Бергмана) или уменьшение придатков (конечностей, ушей, хвоста; правило Аллена) от теплых районов к холодным.